Systematic Paleontology
For the present classification, we follow that by Pteridophyte Phylogeny Group, PPG I [
2].
Class: Polypodiopsida Cronquist, Takhtajan & Zimmermann 1966
Order: Polypodiales Link 1833
Suborder: Dennstaedtiineae Schwarstburd & Hovenkamp 2016
Family: Dennstaedtiaceae Lotsy 1909
Genus:Krameropteris Schneid, Schmidt & Heinrichs 2016
Holotype: Specimen No. PB203297, a pinnule inclusion in Myanmar amber
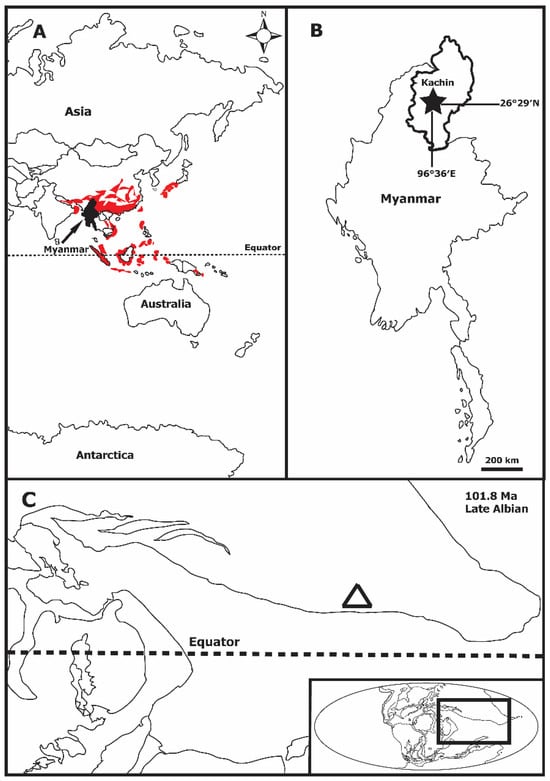
Type locality: Amber mines near Tanai, Ledo Road, 105 km northwest of Myitkyina, Kachin State, Myanmar (26°20′N, 96°36′E). This site is within the Hukawng Basin, which is composed of folded sedimentary (volcanic) rocks of the Cretaceous and Cenozoic Ages.
Type horizon: Myanmar amber, lowest Upper Cretaceous, lowest Cenomanian, absolute age 98.79 ± 0.62 million years ago, established by U-Pb dating of zircons from the rind of the unprocessed amber.
Etymology: The specific epithet calophyllum is proposed to represent the beautiful and well-preserved pinnae fragment. In Old Greek, kallos means “beauty”, while phyllum denotes “leaf”.
Repository: The holotype is deposited in the Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences (abbreviation of NIGPAS), Nanjing, China.
Diagnosis: Fern pinnule pinnate with free branched veins, not reaching the segment margin, tips swollen. Pinnules hypostomatic, short multicellular hairs along pinnule margins, orbicular simultaneous maturity exindusiate sori at ends of veins, polypod sporangia, and trilete tetrahedral–globose spores.
Description: The amber inclusion consists of a single fertile pinnule fragment, measuring 6.4 mm long and 5.2 mm wide. The pinnule is pinnately lobed. The lobes are elliptical with acute apices and slightly thickened margins. The lobes near the apex of the pinnule gradually transition into slightly pinnatifid or crenate forms (
Figure 2A). The venation of the pinnule is pinnate; lateral veins branch 1–3 times with free veinlets. The veinlets terminate at thickened ends located ca. 0.2–0.4 mm from the margins. The sori are abaxial and exindusiate and occur singly, with simultaneous maturity (
Figure 3C,D). They are orbicular, with a maximum diameter of 1.2 mm, and typically one to two sori per pinnule lobe. The more apical pinnule lobes tend to bear a single sorus, while the larger, more basal lobes may contain up to two sori. The sori are submarginal, terminating at the tips of the veinlets, and are located approximately 0.3–1.1 mm from the margins (
Figure 2A). Each sorus contains a somewhat raised receptacle and lacks paraphyses (
Figure 3C,D). Each sorus contains up to 42 mature polypod sporangia, which are nearly globose, measuring 157.3 µm in length and 142.6 µm in width (average of 13 sporangia). The sporangia have stalks with three rows (
Figure 3A), and the annulus is vertical and broken at the stalk, with ~21 darkened and thickened annulus cells (
Figure 3A,B). Some sporangia contain trilete spores, which are tetrahedral–globose in shape, with a conspicuously ornamented perine covered with sparse tubercles and ridges (
Figure 4A,B). Dispersed trilete spores are also present on the abaxial surface of the pinnule (
Figure 4C), with an average diameter of 26–31 μm (measured from seven spores) both in sporangia and on the pinnule abaxial surface. The epidermal cells of the pinnule have undulating or waved anticlinal walls (
Figure 5A). The stomata are hypostomatic and anomocytic, without subsidiary cells, and the guard cells measure 25–40 µm in length (
Figure 3A and
Figure 5B). Short, multicellular hairs, ranging from 18–28 µm in length, are also present along the pinnule margins (
Figure 5C).
Remarks: The fossil represents one terminal fragment of a fertile pinna, containing six sori with developing sporangia and spores. Syninclusions include the remains of being attacked by invertebrates, as well as mites on the pinnule abaxial surface and between the sporangia (
Figure 2). Extant species of
Monachosorum (Dennstaedtiaceae) share similarities with the
Krameropteris fossils in terms of trilete spores, exindusiate sori, and the simultaneous maturity of polypod sporangia. Except for
M. maximowizcii [
29], which differs, the remaining
Monachosorum species have pinnules resembling those of the fossil. Given the uncertainties regarding the evolution of these characteristics, the fossil may not represent a member of
Monachosorum stem lineage, as suggested by Schneider et al. [
5], but could instead be more appropriately placed within the extinct genus
Krameripteris.