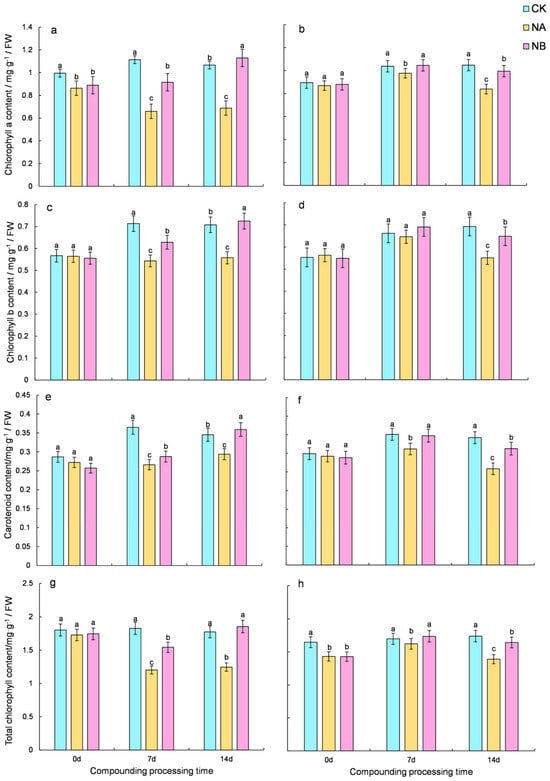
2.1. Effects of Brassinosteroids on Photosynthetic Pigments Under Salt Stress in C. sinensis
To evaluate the impact of 0.2 mg/L BR on levels of photosynthetic pigments, specifically chlorophyll and carotenoids, under salt stress, this study monitored the changes in these pigments. Throughout the treatment period, the two tea varieties exhibited comparable trends in photosynthetic contents (
Figure 1). Under salt stress (NA), the photosynthetic pigment contents decreased. In contrast, the application of 0.2 mg/L BR (NB) effectively increased the pigment levels, approaching those under normal conditions (CK).
In the salinity treatment (NA), the photosynthetic pigment contents of ‘FD’ and ‘CC’ varieties were markedly reduced. Compared with the normal conditions (CK), the total chlorophyll content of the two tea varieties decreased by 34.17% and 4.47% at 7 days and 29.8% and 20.08% at 14 days, respectively. Upon NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in total chlorophyll content compared to that under salt stress at 7 days and 14 days, with increases of 28.33% and 7.01% for ‘FD’ and 48.86% and 18.21% for ‘CC’, respectively (
Figure 1g,h). Compared with the normal conditions (CK), the carotenoids content of the two tea varieties decreased by 27.11% and 11.13% at 7 days and 14.95% and 24.43% at 14 days, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in carotenoid content compared to that under salt stress at 7 days and 14 days, with increases of 8.14% and 11.43% for ‘FD’ and 22.26% and 20.96% for ‘CC’, respectively (
Figure 1e,f).
It was observed that salt stress reduced the photosynthetic pigments in ‘FD’, and the application of 0.2 mg/L BR mitigated the loss of photosynthetic pigments in ‘FD’. The higher chlorophyll content in ‘CC’ than in ‘FD’ suggested that ‘CC’ was more salt-tolerant.
2.3. Effect of Brassinosteroids on the Enzyme Antioxidant System of Camellia Sinensis Under Salt Stress
As illustrated in
Figure 3, salt stress induces a decline in the activity of antioxidant enzymes (POD, CAT, and SOD) in tea plants. However, following BR treatment, the overall activity of antioxidant enzymes in tea plants exhibits an upward trend. Compared with the normal conditions (CK), the SOD activity of the two tea varieties decreased by 24.46% and 20.65% at 7 days and 33.78% and 27.14% at 14 days, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in SOD activity compared to that under salt stress at 7 days and 14 days, with increases of 3.50% and 1.52% for ‘FD’ and 35.29% and 20.21% for ‘CC’, respectively. (
Figure 3a,b). Compared with the normal conditions (CK), the POD activity of the two tea varieties decreased by 40.29% and 51.88% at 7 days and 49.65% and 61.09% at 14 days, respectively. Upon NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in POD activity compared to that under salt stress at 7 days and 14 days, with increases of 14.88% and 24.82% for ‘FD’ and 56.79% and 83.60% for ‘CC’, respectively. (
Figure 3c,d). Compared with the normal conditions (CK), the CAT activity of the two tea varieties decreased by 24.84% and 40.53% at 7 days and 16.28% and 29.08% at 14 days, respectively. Following NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in CAT activity compared to that under salt stress at 7 days and 14 days, with increases of 6.42% and 12.10% for ‘FD’ and 4.44% and 15.50% for ‘CC’, respectively (
Figure 3e,f). Overall, the activity of antioxidant enzymes in tea plants declined under salt stress, whereas it increased after BR spraying, yet failed to revert to the normal level.
2.4. Changes in Contents of Soluble Sugars, Soluble Proteins, and Proline
Exogenous spraying of 0.2 mg/L BR under salt stress conditions led to an increase in the levels of soluble sugars, soluble proteins, and proline in both varieties. Relative to the normal control (CK), the soluble sugar content of the two tea varieties increased by 16.27% and 32.65% at 7 days and 22.14% and 52.23% at 14 days, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in soluble sugar content compared to that under salt stress at 7th day and 14th day, with increases of 10.83% and 13.20% for ‘FD’ and 25.00% and 12.76% for ‘CC’, respectively (
Figure 4a,b). Compared to the normal conditions (CK), the soluble protein content of the two tea varieties increased by 66.12% and 55.48% at 7 days and 74.52% and 71.04% at 14 days, respectively. Upon NB treatment, both ‘FD’ and ‘CC’ varieties demonstrated a significant increase in soluble protein content compared to that under salt stress at 7 days and 14 days, with increases of 13.10% and 5.34% for ‘FD’ and 27.93% and 21.18% for ‘CC’, respectively (
Figure 4c,d). Compared with the normal control (CK), the proline content of the two tea varieties increased by 39.17% and 95.90% at 7 days and 71.02% and 245.30% at 14 days, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in proline content compared to that under salt stress at 7 days and 14 days, with increases of 7.68% and 38.97% for ‘FD’ and 26.43% and 32.66% for ‘CC’, respectively (
Figure 4e,f). However, under salt stress, the contents of soluble sugar, soluble protein, and proline in ‘CC’ were higher than those of ‘FD’ under salt stress, suggesting that NA promotes the accumulation of these substances in tea plants. Exogenous spraying of 0.2 mg/L BR has a beneficial effect on tea plants under salt stress, and this effect becomes more pronounced over time.
2.5. The Effect of Exogenous Brassinosteroids (BRs) on the Expression of Genes Related to the Proline Metabolism Pathway in Tea Plants Under Salt Stress
Analysis of the physiological data from this experiment revealed that proline levels were significantly affected by NA and NB treatment. By referring to the Camellia Sinensis ‘Shuchazao’ tea plant genome database, six genes associated with the proline metabolism pathway were identified, namely CsProDH–A, CsProDH–B, CsP5CS–A, CsP5CS–B, CsOAT–A, and CsOAT–B.
The
CsProDH gene encodes proline dehydrogenase (
ProDH). Relative to the normal control (CK), the expression of
CsProDH–
A in the two tea varieties was downregulated by 61.20% and 37.14% at 7 days and 30.39% and 31.97% at 14 days, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant upregulation in
CsProDH–
A compared to that under salt stress at 7 days and 14 days, with upregulations of 126.33% and 28.21% for ‘FD’ and 85.32% and 26.26% for ‘CC’, respectively (
Figure 5a,b). Compared with the normal conditions (CK), the expression of
CsProDH–
B in the two tea varieties was downregulated by 33.65% and 42.69% at 7 days and 10.70% and 26.53% at 14 days, respectively. The extent of downregulation of
CsProDH–
B was greater in ‘CC’ than in ‘FD’. However, after NB treatment, the expression of
CsProDH–
B in the ‘FD’ variety showed a trend of initial upregulation followed by downregulation on the 7th and 14th days compared to that under salt stress, with rates of 10.02% and 1.84%, respectively. In contrast, the expression of
CsProDH–
B in the ‘CC’ variety exhibited a trend of first downregulation and then upregulation under salt stress on the 7th and 14th days, with values of 23.97% and 18.33%, respectively (
Figure 5c,d). These results suggest that the expression of the
CsProDH gene regulates the degradation of proline.
The protein encoded by the
CsP5CS gene is pyrroline-5-carboxylate synthetase (P5CS), a key enzyme in proline synthesis in plants. Relative to the normal control (CK), the expression of
CsP5CS–
A in the ‘FD’ variety was upregulated by 60.80% and 21.18% at 7 days and 14 days, respectively. Under salt stress, the expression of
CsP5CS–
A in the ‘CC’ variety showed a trend of initial downregulation followed by upregulation on the 7th and 14 days, with rates of 25.89% and 64.05%, respectively. After NB treatment, the expression of
CsP5CS–
A in the ‘FD’ variety was upregulated by 17.80% and 4.64% at 7 days and 14 days, respectively. In contrast, the expression of
CsP5CS–
A in the ‘CC’ variety exhibited a trend of first upregulation and then downregulation under salt stress on at 7 days and 14 days, with rates of 15.77% and 8.33%, respectively (
Figure 5e,f). Compared to the normal control (CK), the expression of
CsP5CS–
B in the two tea varieties was upregulated by 126.92% and 380.87% at 7 days and 73.16% and 316.82% at 14 days, respectively. The extent of upregulation of
CsP5CS–
B was greater in ‘CC’ than in ‘FD’. After NB treatment, the expression of
CsP5CS–
B in the ‘FD’ variety was upregulated by 21.40% and 14.88% at 7 days and 14 days, respectively. In contrast, the expression of
CsP5CS–
B in the ‘CC’ variety exhibited a trend of first downregulation and then upregulation under salt stress on the 7th and 14th day, with rates of 45.43% and 6.54%, respectively (
Figure 5g,h).
Relative to the normal control (CK), the expression of
CsOAT–
A in the ‘FD’ variety was upregulated by 16.17% at 14 days. In the ‘CC’ variety, it was upregulated by 16.05% and 11.62% at 7 days and 14 days, respectively. After NB treatment, the expression of
CsOAT–
A in the ‘FD’ variety was upregulated by 13.57% and 40.56% at 7 days and 14 days, respectively. In contrast, the expression of
CsOAT–
A in the ‘CC’ variety exhibited a trend of first upregulation and then downregulation under salt stress on the 7th and 14th day, with rates of 8.53% and 8.36%, respectively (
Figure 5i,j). Relative to the normal control (CK), the expression of
CsOAT–
B in the two tea varieties was upregulated by 39.79% and 32.90% at 7 days and 22.03% and 12.10% at 14 days, respectively. After NB treatment, the expression of
CsOAT–
B in the two tea varieties showed a trend of initial downregulation followed by upregulation under salt stress on the 7th and 14th day, with rates of 0.15% and 8.76% and 46.85% and 7.20%, respectively (
Figure 5k,l).
Gene expression analysis revealed that the CsP5CS gene was initially downregulated and subsequently upregulated under salt stress, whereas the CsProDH gene was downregulated throughout. Regarding CsOAT, its expression patterns varied among different varieties under salt stress. These findings suggest that the upregulation of CsP5CS gene expression facilitates proline synthesis. Moreover, exogenous BR treatment promotes CsOAT gene expression and activates the ornithine synthesis pathway, thereby enhancing proline accumulation.
2.6. The Effect of Exogenous Brassinosteroids (BRs) on the Expression of Genes Related to the Flavonoid Synthesis Pathway in Tea Plants Under Salt Stress
By referring to the ‘Shuchazao’ tea plant genome database, sixteen genes associated with the flavonoid metabolism pathway were identified, namely CsPAL, CsC4Ha, CsC4Hb, Cs4CL, CsCHSa, CsCHSb, CsF3Ha, CsF3Hb, CsF3′5′Ha, CsF3′5′Hb, CsDFRa, CsDFRb, CsLAR, CsANRa, CsANRb, and CsANS. Their expression characteristics were subsequently analyzed using qRT-PCR.
Under salt stress treatment, all genes associated with the flavonoid synthesis pathway in both varieties exhibited an upregulation trend (
Figure 6). In comparison to the salt stress treatment group, the BR spraying treatment groups also showed an upregulation trend, suggesting that BR spraying induced the expression of genes related to the flavonoid synthesis pathway, leading to an increased effect.
2.7. The Effect of Exogenous Brassinosteroids (BRs) on the Expression of Genes Related to the Theanine Synthesis Pathway in Tea Plants Under Salt Stress
Four genes related to the theanine synthesis pathway, namely CsTSa, CsTSb, CsGSa, and CsGSb, were identified from the ‘Shuchazao’ tea plant genome database, and their expression characteristics were analyzed using qRT-PCR.
As depicted in
Figure 7, genes related to the theanine synthesis pathway generally exhibited an upward trend. Relative to the control (CK), both ‘FD’ and ‘CC’ varieties showed a significant increase in
CsTSa expression at 7th day and 14th day, with upregulations of 15.52% and 26.48% for ‘FD’ and 40.07% and 39.90% for ‘CC’, respectively. After NB treatment, the expression of
CsTSa in ‘FD’ was upregulated by 109.01% at 7th day and tended to return to the level of CK at 14th day. For ‘CC’,
CsTSa was upregulated by 86.00% and 23.02% at 7th day and 14th day, respectively (
Figure 7a,b). Compared to the control (CK), the expression of
CsTSb in the two tea varieties was upregulated by 59.37% and 37.23% at 7 days and 96.13% and 49.09% at 14 days, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant increase in
CsTSb expression at 7th day and 14th day, with upregulations of 27.07% and 3.58% for ‘FD’ and 53.30% and 59.36% for ‘CC’, respectively (
Figure 7c,d).
Relative to the control (CK), both ‘FD’ and ‘CC’ varieties showed a significant increase in
CsGSa expression at 7th day and 14th day, with upregulations of 73.26% and 32.02% for ‘FD’ and 53.87% and 17.83% for ‘CC’, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties exhibited a significant increase in
CsGSa expression at 7th day and 14th day, with upregulations of 17.17% and 39.01% for ‘FD’ and 46.70% and 22.45% for ‘CC’, respectively (
Figure 7e,f). Compared to the control (CK), the expression of
CsGSb in the two tea varieties was upregulated by 12.94% and 41.21% at 7th day and 76.27% and 98.05% at 14th day, respectively. After NB treatment, the expression of
CsGSb in the two tea varieties was upregulated by 59.02% and 5.94% at 7th day and 50.34% and 31.52% at 14th day, respectively (
Figure 7g,h).
The CsTS gene is closely associated with theanine synthesis in tea plants. When tea plants are exposed to salt stress, CsTS is induced to express, potentially leading to increased theanine synthesis. Overall, both CsTS and CsGS genes were upregulated under salt stress. After BR spraying, an enhancing effect was observed, and their expression levels gradually returned to normal at 14th day.
2.8. Genes Related to Stress
Four adversity marker genes, namely CsRD29A, CsRD29B, CsRD20, and CsADH1, were identified from the genome database of Camellia sinensis ‘Shuchazao’, and their expression characteristics were analyzed using qRT-PCR.
As shown in
Figure 8, the expression of genes associated with stress significantly increased upon exposure to salt stress (NA) and compound treatment (NB). Relative to the normal control (CK), both ‘FD’ and ‘CC’ varieties showed a significant upregulation in
CsRD29B expression at 7th day and 14th day, with upregulations of 17.25% and 41.38% for ‘FD’ and 41.42% and 10.72% for ‘CC’, respectively. After NB treatment, the expression of
CsRD29B in the two tea varieties was upregulated by 9.17% and 0.39% at 7th day and 7.17% and 6.00% at 14th day, respectively (
Figure 8a,b). Compared to the control (CK), the expression of
CsRD29A in the two tea varieties was upregulated by 148.27% and 24.05% at 7th day and 12.89% and 100.01% at 14th day, respectively. After NB treatment, both ‘FD’ and ‘CC’ varieties showed a significant upregulation in
CsRD29A expression at 7th day and 14th day, with upregulations of 86.59% and 61.60% for ‘FD’ and 158.10% and 19.75% for ‘CC’, respectively (
Figure 8c,d). The extent of
CsRD29A upregulation was higher in ‘FD’ than in ‘CC’. Generally, under the composite treatment,
CsRD29A levels in both varieties were significantly higher than those in other treatment groups, implying that this gene plays a crucial role in the response to salt stress when sprayed with 0.2 mg/L BR. Additionally, the expression level of
CsRD29A in the combined treatment was higher than that under salt stress alone, suggesting that BR can enhance the plant’s response to salt stress. The
CsRD29B gene in ‘CC’ also responds to salt stress, albeit less prominently than
CsRD29A.
Relative to the control (CK), both ‘FD’ and ‘CC’ varieties showed a significant upregulation in
CsRD20 expression at 7th day and 14th day, with upregulations of 463.94% and 165.92% for ‘FD’ and 748.97% and 193.37% for ‘CC’, respectively. The extent of
CsRD20 upregulation was higher in ‘FD’ than in ‘CC’. After NB treatment, the expression of
CsRD20 in ‘FD’ was downregulated by 32.39% and 37.48% at 7th day and 14th day, respectively. In contrast, the expression of
CsRD20 in ‘CC’ was upregulated by 2.69% and 486.53% at 7th day and 14th day, respectively (
Figure 8e,f). Overall,
CsRD20 acts as a rapid early responder. In ‘FD’ plants, exogenous BR treatment alleviates its high expression, whereas in ‘CC’ plants, the gene response is slower. Additionally, the
CsADH1 gene exhibits differential expression patterns across varieties.
Relative to the control (CK), both ‘FD’ and ‘CC’ varieties exhibited a significant upregulation in
CsADH1 expression at 7th day and 14th day, with upregulations of 114.82% and 315.43% for ‘FD’ and 8.74% and 272.41% for ‘CC’, respectively. The extent of
CsADH1 upregulation was higher in ‘CC’ than in ‘FD’. After NB treatment, the expression of
CsADH1 in ‘FD’ was downregulated by 12.95% and 7.76% at 7th day and 14th day, respectively, while that in ‘CC’ was upregulated by 4.02% and 25.71%. At the same time points, ‘FD’ and ‘CC’ varieties exhibited a more significant enhancement than CK at 7 d and 14 d in
CsADH1, and expression was upregulated by 114.82% and 315.43% and 8.74% and 272.41%, respectively. The upregulation of
CsADH1 in ‘CC’ is higher than that in ‘FD’. Under NB treatment,
CsADH1 expression in the ‘FD’ variety at 7 d and 14 d was downregulated by 12.95% and 7.76%.
CsADH1 expression in the ‘CC’ variety at 7 d and 14 d was upregulated by 4.02% and 25.71% (
Figure 8g,h).