1. Introduction
Cardiovascular disease (CVD) is now the major cause contributing to disability and premature death globally. According to the report of Global Burden of Diseases, Injuries, and Risk Factor Study 2015 and the WHO, CVD affected an estimated 422.7 million people and caused an estimated 17.9 million deaths worldwide in 2019, comprising 32% of all deaths worldwide [
1,
2]. By 2030, the annual cardiovascular disease mortality is expected to reach 23.6 million. The large and progressively increasing burden of CVDs on people, families, and healthcare systems is an indicative of an urge to conduct the research on atherosclerotic diseases and the adoption of preventive measures [
3,
4].
Research is always focused on identifying reliable biomarkers for CVD. Numerous biomarkers linked to the development of CVD include a series of events initiated by inflammation and endothelial damage [
5]. Potential biomarkers for CVD prediction include elevated levels of homocysteine, the erythrocyte sedimentation rate, and C-reactive protein [
6].
Homocysteine (Hcy) is a naturally occurring non-proteinogenic sulfur-containing α-amino acid in human body that functions as a key intermediate in the biosynthesis of methionine and cysteine [
7,
8]. Methionine undergoes methylation by methionine adenosyltransferase (MATs) (also known as
S-Adenosylmethioninesynthetase) to form
S-Adenosylmethionine (SAM), which further irreversibly converts to
S-adenosylhomocysteine (SAH), and SAH is further converted to Hcy. The metabolism of Hcy stands at the intersection of two pathways. The first pathway involves the remethylation to methionine, which requires folate and vitamin B
12; second, Hcy is trans-sulfurated to cystathionine to cysteine, which requires vitamin B
6 [
7,
9]. The blockage of either the cysteine or methionine pathway may increase the Hcy levels, leading to hyperhomocysteinemia (hHcy). An abnormally high level of Hcy in the serum is major risk factor for the development of a wide range of disorders such as thrombosis, neuropsychiatric illness, and fractures [
10]. Significant associations exist between hHcy and cardiovascular disease and its repercussions such as cardiac arrests and strokes. Additionally, it has been discovered to be linked to microalbuminuria, a significant predictor of future cardiovascular disease [
11,
12].
Thus, the hHcy may be involved in oxidative stress, inflammation, and ischaemic alterations [
13]. The blood artery structure and function are modified by hHcy, and oxidative stress seems to be the main mechanism causing these changes [
14,
15]. Cardiovascular illnesses are caused by the reactive oxygen species that are produced, which also promote the production of superoxide radicals, lipid peroxidation, and protein oxidation [
16,
17]. Studies have demonstrated a reduction in levels of homocysteine with folate, which is the primary vitamin utilized to lower homocysteine levels. A greater reduction in homocysteine levels can be achieved with vitamin B12 plus folate than with folate alone. Other than vitamin supplements, there are no other treatments available. Strong potential exists for plants high in phenolic acids to serve as major sources of naturally occurring antioxidants [
18]. Numerous traditional plants, primarily ferulic acid, are excellent sources of phenolic acid. Ferulic acid, also known as 3-(4-hydroxy-3-methoxy-phenyl) prop-2-enoic acid, is a common phenolic acid that may be found in a wide variety of plants [
19]. It is abundant in many staple foods, including citrus fruits, whole grains, spinach, broccoli, and grain bran, and it is found in plant tissues [
20]. In China, it is used clinically to treat hypertensive disorders and angina pectoris [
21]. Furthermore, the evidence from epidemiological research suggests that consuming whole grains high in FA lowers the chance of developing chronic illnesses, such as cardiovascular disease [
20]. Its therapeutic effects are multifaceted and include antioxidant [
22,
23], anti-hypertensive [
24], anti-inflammatory [
25], neuroprotective [
26], and cardioprotective [
27] properties. The present investigation was conducted with the aim of assessing the impact of FA on hHcy in Wistar rats induced by L-methionine.
3. Discussion
This epidemiological study indicates that a mild elevation of plasma Hcy concentration is an independent risk for atherothrombotic diseases [
28,
29]. In the general population, increased Hcy is common with individual suffering from coronary, cerebro-vascular, and peripheral atherosclerotic diseases [
30,
31]. A common reason for increase Hcy is the inadequate concentration of vitamins required for Hcy metabolism such as folic acid and vitamin B12 and B6 [
32,
33]. In addition to low supply, an increased requirement for folic acid or the interference of drugs with folate metabolism can cause a folate deficiency. Drugs like aminopterin (immunosuppressive), methotrexate (anti-cancer), pyrimethamine (anti-protozoal), trimethoprim (urinary tract infection), and triamterene (sodium channel blocker) affect the folate metabolism [
34,
35]. A higher concentration of Hcy is also common in pregnant women, smokers, alcoholics, and users of contraceptive medicines due to an alteration in the enzymes controlling Hcy metabolism [
36,
37,
38,
39]. Elevated levels of Hcy can modify the morphology of the blood vessel wall, increase platelet activity, and stimulate smooth muscle proliferation and LDL oxidation. Hcy adversely affects the endothelial morphology of arteries and arterioles. hHcy has also been reported to affect the cardiovascular system, thereby leading to cardiovascular diseases [
9].
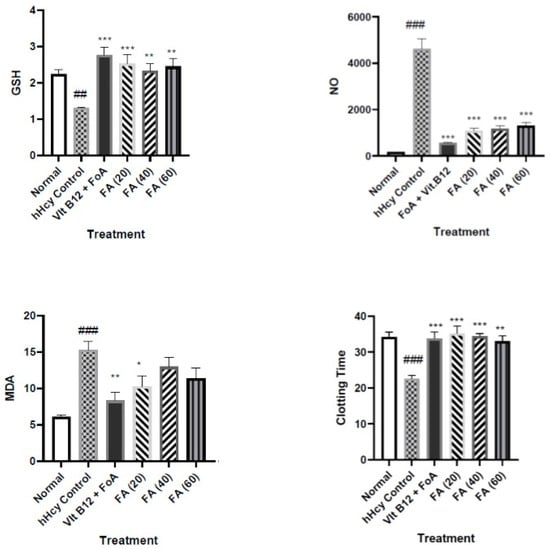
In the present study, we administered methionine (1 gm/kg) for 36 days to Wistar rats to induce hHcY. It was observed that Hcy was increased five-fold in hHcy-controlled animals, while FA was able to reduce it significantly. The effects were comparable to the standard, vit. B12, and FoA group. Previously, we have reported the cardioprotective activity of FA in the abdominal aortic banding (AAB) model of myocardial infarction [
40] and in doxorubicin-induced cardiac toxicity [
41]. The potential cardioprotective activity of FA is attributed to its potent antioxidant profile [
42]. FA is a derivative of oryzanol, which is now intentionally added to edible oils and is a key component of rice bran oil. The oryzanol component of rice bran oil is a fraction containing FA that accounts for 80% of oryzanol. Oryzanol is reported to be a potent antioxidant due to its FA content being higher than vitamin E [
43]. As previously mentioned, FA is known to reduce cholesterol, oxidized LDL-C, and oxidative stress and can therefore improve the lipid profile and blood vessel activity, hence reducing cardiovascular disease risk factors [
20,
21,
44,
45]. Ferulic acid is abundantly found in fruits and vegetables, such as sweet corn, tomatoes, and in rice grain, thus the intake of FA-rich food can help to alleviate the risk of hHcy.
Although hyperhomocysteinemia is commonly caused by vitamin deficiency, other causes may include the dysfunction of enzymes and cofactors associated with the process of homocysteine biosynthesis, excessive methionine intake, certain diseases, and the side effects of some drugs. Persistent hHcy leads to increased serum cholesterol, increased LDL, and reduced HDL cholesterol [
46,
47], which are similar findings we have obtained. Hyperhomocysteinemia produces oxidative stress, which contribute to atherosclerosis. The free oxygen radicals forms oxidized LDL in the endothelial tissues of blood vessels, and oxidized LDL promotes the formation of foam cells, leading to atherosclerotic lesions. FA has been reported to lower the plasma levels of total cholesterol, VLDL, and LDL [
48]. It has also been reported to inhibit hepatic HMG-CoA reductase, the rate-limiting enzyme in cholesterol biosynthesis and acyl-CoA cholesterol esterifying enzyme in tissue [
49]; our results are in agreement with the reported studies. The histopathology of the aorta in the present study confirms these findings. The histopathology indicated the presence of atherosclerotic lesions in the hHcy group while it was absent in treatment groups. Although betaine, a byproduct of choline oxidation, is known to lower hHcy, it also has a negative impact on hyperlipidemia. As a result, FA is a superior option for the treatment of hHcy due to its homocysteine-lowering property and its hypolipidemic effect.
Oxidative stress brought on by hHcy may also result in reduced glutathione, lipid peroxidation (MDA), and an increase in total proteins. Our findings are close to those of the earlier research [
40]. Peroxynitrite is formed when nitric oxide and superoxide anion, which are generated during hHcy-induced oxidative stress [
50]. We measured the amount of NO metabolites and discovered a substantial rise in hHcy, which may be related to nitrosive stress. We speculate that both peroxide and peroxy nitrite production are responsible for the oxidative stress generated in hHcy. As previously noted, FA has a strong effect on inhibiting the development of both nitrite and oxide [
51]. According to Woo et al. (2006), hHcy induces oxidative stress in the liver, which might raise ALT and AST levels [
52]. Based on AST and ALT data, FA, a free radical scavenger, reduced liver damage. As previously reported, we also found a prominent effect at the lower dose of FA (at 20 mg/kg, p.o.) [
53]. The reason could be attributed to the formation of undesired metabolites of FA (as given in earlier reports) or it may be due to the prolonged use (36 days of administration) of FA leading to the achievement of the steady-state concentration.
Increased oxidative stress causes damage to the heart’s cells in hHcy, which raises the blood levels of LDH and CK-MB. In the current investigation, hHcy’s detrimental effect on cardiac cells has been found to raise the serum levels of CK-MB and LDH, whereas the FA treatment significantly decreased both of these levels, indicating its cardioprotective action. It has been reported that an increased level of Hcy is an independent risk factor for premature cardiovascular diseases [
9]. An elevated Hcy concentration leads to the constriction of blood vessels, resulting in atherogenesis and thrombosis. The present study demonstrated the influence of hHcy on hemodynamic and electrocardiographic parameters, but we found that hHcy did not increase blood pressure and did not affect ECG. However, the increased treatment days of methionine can cause an alteration in BP and ECG; this possibility cannot be denied. It has been reported that a higher plasma total Hcy concentration is linked to enhanced platelet aggregation and thromboxane synthesis, which is a key factor for blood clotting [
54]. In our study, we have observed similar findings; the blood clotting time has been increased due to increased Hcy concentration. FA restored the clotting time to normal.
FA is a powerful antioxidant, but our research is the first to suggest that it affects the homocysteine and methionine cycle. The figure illustrates the possible mechanism of action (
Figure 4). As previously noted, ferulic acid may increase the effectiveness of SAM for the further methylation of biomolecules including proteins, nucleic acid, etc., thereby lowering the production of homocysteine [
55].
The primary function of MAT, a rate-limiting enzyme in the methionine cycle, is to catalyze the conversion of methionine and adenosine triphosphate (ATP) into S-adenosylmethionine (SAM), and SAM serves as the methyl donor in almost all methylation reactions within the body. SAM is irreversibly transformed into the metabolic precursor of homocysteine, S-adenosylhomocysteine (SAH). Hence, the modulation of SAM levels through MAT inhibition could be a possible mechanism of action, as shown by the Hcy level in the treatment group compared to the diseased group in the present study, and is further supported by the docking results. Thus, docking and biochemical studies show that FA may increase the activity of the SAM/SAH-dependent methylation of other proteins and allows progress towards Hcy formation. However, it is necessary to measure the levels of SAM and SAH to further confirm this mechanism.
4. Materials and Methods
4.1. Animals
Adult male Wistar rats (160–200 g) were obtained from the Institutional Animal House of SIOP (CPCSEA/1139/a/07) and housed in groups of six animals per cage at standard laboratory conditions with a temperature of 25 ± 1 °C, relative humidity of 45–55%, and 12:12 h dark and light cycle. Animals had free access to food (standard chow pellet, Nutrivet Life Sciences, Pune, India) and water ad libitum. The experimental protocol was approved by the Institutional Animal Ethics Committee (IAEC) of Sinhgad Institute of Pharmacy, Narhe, Pune, India, constituted according to guidelines of Committee for the Purpose of Control and Supervision of Experiment on Animals (reg. no. SIOP/IAEC/2014/22).
4.2. Chemicals
Ferulic Acid (Sigma Aldrich, St. Louis, MO, USA), L-Methionine (Loba Chemicals, Mumbai, India), and reduced glutathione (GSH) were obtained from LobaChemie Pvt. Ltd. (Mumbai, India). CK-MB, alanine transferase (ALT) and aspartate transferase (AST) kits (Pathozyme Diagnostics, Kolhapur, Maharashtra, India), and lactate dehydrogenase (LDH) kits (Bio-Lab Diagnostics, Mumbai, India), total cholesterol (Span Diagnostics Ltd., Pune, India), and Rat Hcy ELISA (Blue Gene Biotech Co., Ltd., Shanghai, China) were purchased.
4.3. Experimental Protocol for L-Methionine-Induced hHcy in Rats
Thirty-six healthy adult male Wistar rats (160–200 g) were used for this study. The animals were randomly divided into six groups (
n = 6). Group I: normal was given distilled water. Group II: hHcy control (disease induced) was treated with L-methionine (1 gm/kg). Group III: standard group received a standard treatment of vit. B12 (15 mg/kg) + folic acid (FoA); 70 mg/kg). Group IV, V, and VI were treated with test drug, FA (20/40/60 mg/kg, respectively). All the treatments were given orally for 36 days simultaneously, and Met (1 gm/kg. p.o.) administration was performed prior to 1 h in all the groups except normal control. On day 28, blood was removed using retroorbital plexus method and serum was separated. Levels of Hcy, cholesterol, LDH, CK-MB, ALT, AST were determined in serum. On day 36, blood pressure, both systolic and diastolic, heart rate, and ECG were determined. The animals were then sacrificed humanely. Liver was carefully separated and used for the estimation of MDA, GSH, and nitric oxide (NO). Heart and abdominal aorta were separated for histopathological studies [
56,
57].
4.4. Pharmacological Evaluation: Assessment of Hemodynamic and Electrocardiographic Parameters
On the last day of study, animals were anesthetized with urethane (1.25 g/kg, i.p.). The right carotid artery of each rat was cannulated for the measurement of systolic blood pressure (SBP) and diastolic blood pressure (DBP). The cannula was filled with heparinized saline and connected to pressure transducer. After 30 min of stabilization, hemodynamic parameters were recorded by eight channel recorder Power lab (AD Instruments Pty Ltd. with LABCHART Pro software version 7, Bella Vista, NSW, Australia). For the measurement of the electrocardiograph, the leads were placed on the right foreleg (positive electrode), left foreleg (negative electrode), and right hind leg (neutral electrode) [
58,
59].
4.5. Serum Parameters
Blood was removed; serum was separated and kept at −20 °C for the estimation of CKMB, LDH, AST, ALT, cholesterol, Hcy, and total protein level. The serum CK-MB, LDH, AST, and ALT activity was measured using reagent kits and an automated biochemistry analyzer (Mindray BC-2800, Shenzhen, China) [
60]. The serum Hcy level was measured using an ELISA kit (Blue Gene Biotech Co., Ltd., Shanghai, China) [
61].
4.6. Tissue Parameters
Liver was quickly transferred to ice-cold phosphate-buffered saline (pH 7.4). It was blotted free of blood and tissue fluids. The hearts were cross-chopped with a surgical scalpel into fine slices, suspended in chilled 0.25 M sucrose solution, and quickly blotted on a filter paper. The tissues were then minced and homogenized in chilled tris hydrochloride buffer (10 mM, pH 7.4) to a concentration of 10% w/v. Prolonged homogenization under hypotonic condition was designed to disrupt, as far as possible, the structure of the cells so as to release soluble proteins. The homogenate was centrifuged at 7000 rpm at 25 min using high-speed cooling centrifuge (Remi R-8C, Remi laboratory Instruments, Mumbai, India). The clear supernatant was used for the estimation of tissue parameters.
4.7. GSH Estimation
Equal volumes of liver tissue homogenate (supernatant) and 20% trichloroacetic acid (TCA) were mixed. This precipitated fraction was centrifuged, and to 0.25 mL of supernatant, 2.0 mL of DTNB reagent was added. Final volume was made up to 3.0 mL with phosphate buffer. The color developed was read at 412 nM against the reagent blank. The amount of reduced glutathione was expressed as mg of GSH/g of wet tissue [
62].
4.8. Malondialdehyde (MDA) Estimation
An amount of 0.2 mL of liver tissue homogenate was added in each test tube; 0.2 mL of reagent-A, 1.5 mL of B, and 1.5 mL of C were added in each test tube mixture, which finally made up a total of 4 mL with water and was then heated at 95 °C for 60 min in oil bath. Then, solution was cooled under tap water, and 1 mL water and 5 mL of Reagent D were added in each test tube. These mixtures were vortexed and centrifuged at 2200 g for 5 m. Organic layer (upper layer) was discarded and aqueous layer (lower layer) was used to determine the absorbance colorimetrically at 532 nm against reagent blank [
62].
4.9. Nitric Oxide (NO) Estimation
An amount of 0.1 mL of tissue homogenate was added to each test tube containing 10mL, followed by the addition of 500 μL of Griess reagent to each test tube; the absorbance was determined at 546 nm. Then, concentration was measured against reagent blank by interpolating absorbance on standard curve and the values were expressed as µg/mL [
63,
64].
4.10. Histopathology
The animals were euthanized and the abdominal aorta was removed and placed in 10% formalin solution. The organ specimens were subjected to dehydration with xylene (one hour each) and alcohol of 70, 90, and 100% strength each for two hours. The infiltration and impregnation were carried out by treatment with paraffin wax twice for each time for one hour. Paraffin wax was used to prepare paraffin L molds. Specimens were cut into sections of 3–5 mm thickness and stained with orcein dye. For 10 randomly chosen microscopic fields (40× magnification) of each left ventricle, Masson’s trichome stained myocardial sections were imaged. This served as an estimate of the atherosclerotic lesions [
41].
4.11. Docking Studies
The docking studies were performed using AutoDock-4.2 software, employing the Lamarckian Genetic Algorithm (LGA) for conformational search. Visualization of the docking results was performed using Discovery Studio Visualizer-2019. The protein models for methionine adenosyltransferase with PDB id-190 was obtained from the protein databank (
https://www.rcsb.org/; accessed on 25 October 2023). The refinement of protein structures was performed through dockprep utility within Chimera-1.17. Ferulic acid structures were minimized through the MMFF94s force field. The grid box was characterized by dimensions of 68 units along each axis (X, Y, Z), with its center located at coordinates (2.054, 36.636, 54.828) for the X, Y, and Z dimensions, respectively, and then the binding energy and conformation was assessed [
65,
66,
67].
4.12. Statistical Analysis
Data for each parameter was analyzed by one-way ANOVA followed by Dunnett’s post hoc test using a graph pad and Prism software, version 5.0, Prism Software Inc., La Jolla, CA, USA.