
1. Introduction
Regions characterized by permafrost cover approximately 22% of the Northern Hemisphere’s terrestrial area [1]. These areas are currently experiencing swift deterioration as a consequence of global warming [2]. The thawing of permafrost can lead to the decomposition of organic materials that have been stored in the soil for extended periods, subsequently releasing various greenhouse gasses such as CO2, CH4, and N2O into the atmosphere. This process contributes to a self-reinforcing loop that exacerbates climate change [3]. In 2023, global greenhouse gas emissions reached a record high of 57.1 billion tons of CO2 Eq [4]. According to the latest data on greenhouse gas release from the permafrost of the Northern Hemisphere, with global warming, permafrost regions have experienced significant warming, with rates 2 to 4 times higher than the global average [5].
The release of greenhouse gasses (GHGs) from permafrost regions essentially results from the microbial utilization of organic matter, and this process is mainly affected by the substrate supply and microbial communities [6]. In areas with a high prevalence of permafrost, the soil’s organic matter has been accumulating gradually over a long period, leading to a nutrient-deficient soil profile. This results in a high carbon-to-nitrogen ratio, which in turn restricts microbial activity during the decomposition of organic matter due to a lack of nitrogen [7]. Consequently, the addition of nitrogen often enhances the breakdown of the soil organic matter (SOM) [8,9,10,11]. An illustrative case is the application of nitrogen fertilizers to an alpine meadow on the northern edge of the Qinghai–Tibet Plateau; this was observed to boost soil microbial activity and elevate CO2 concentrations. However, there was no significant impact on the absorption of CH4 or the emission of N2O [12]. However, it was also found that the high concentration of nitrogen added (6 g nitrogen m−2 a−1) inhibited the emission of CH4 in the boreal peatlands of northeast China [13].
Nitrogen deposition controls greenhouse gas emission processes by regulating the quantity, function, and structure of soil microorganisms [14]. Oligotrophic bacteria have a strong metabolic capacity for refractory organic matter. These species can survive in nutrient-poor environments (high carbon-to-nitrogen ratio) but grow slowly in nutrient-rich conditions in the short term. Eutrophic bacteria tend to utilize readily available organic carbon and have a high nitrogen requirement [15]. Therefore, it is hypothesized that nitrogen deposition can alleviate nitrogen limitations for microorganisms; thus, the relative abundance of eutrophic bacteria declines [16].
The introduction of nitrogen can influence the flux of CO2 by boosting plant growth, which in turn enhances the nutritional composition of plant debris and mitigates the constraints on microbial metabolism caused by nitrogen scarcity [17,18]. In peatland ecosystems, the primary agents responsible for the decomposition of organic matter are bacteria and fungi [19,20]. After the input of nitrogen, soil microorganisms retain more NH4+, and plants mainly use NO3−. Both of these processes increase the size of the soil microbial community and plant root activity, respectively, increasing the emission of CO2 [21].
CH4 is produced by methanogens under anaerobic conditions [22,23]. The introduction of nitrogen can influence the metabolic processes of methanogenic microorganisms [24] and further affect the production of CH4 [25,26]. Previous findings suggest that the addition of nitrogen can inhibit the absorption of CH4 [27] and increase the emission of CH4 [28]. The input of nitrogen can reduce the capacity of soils to absorb methane, primarily by changing the quantity and variety of methanotrophs and methanogens, as well as their community composition. Moreover, a decrease in the soil pH, which is often associated with increased levels of nitrogen, plays a significant role in governing the diversity and composition of these microbial communities [29].
Enhanced emissions of N2O have been linked to elevated rates of nitrification and denitrification. The addition of nitrogen to soil has been shown to boost the emission of N2O, likely due to the proliferation of bacteria responsible for these processes [30,31,32,33]. Nitrobacter, which plays a key role in ecosystem denitrification, facilitates this process, thereby contributing to the greater release of N2O [34].
Various critical genes involved in the biogeochemical cycles of carbon and nitrogen are frequently utilized to evaluate the population and variety of microbes in soil, encompassing those that are vital for soil respiration processes (bacteria and fungi) [35,36], CH4 generation (mcrA), CH4 oxidation (pmoA) [37,38], N2O production (nirS and nirK) [39,40], and nitrogen fixation (nifH) [41,42]. Although these genes have been recognized, their abundance and relationship with greenhouse gas emissions largely remain unclear; this is because many physiochemical variables can also affect greenhouse gas emissions [43]. Overall, nitrogen addition influences greenhouse gas emission patterns by affecting soil microorganisms. Nitrogen application at different levels (low, medium, and high) may have varying impacts on greenhouse gas emissions. Therefore, it is of great significance to study the changes in soil microbial functional genes under different nitrogen application levels and their relationship with greenhouse gas emissions, which may benefit the prediction of emission patterns in the context of future increases in atmospheric nitrogen deposition.
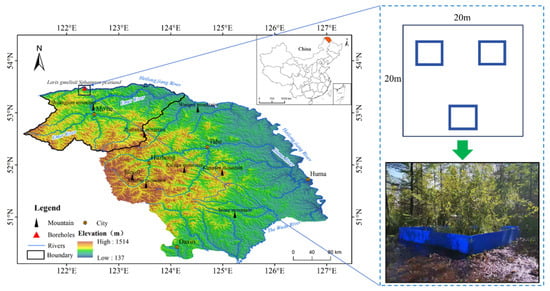
Prolonged cold and wet conditions in northern peatlands have led to the sequestration of over 875 Pg of carbon [44]. The carbon stored in permafrost peatlands accounts for 1/3 of the soil carbon pool in the global terrestrial ecosystem [45,46]. Permafrost peatlands are nutrient-poor, limited by nitrogen, and sensitive to changes in nitrogen levels [47]. The continuous increase in nitrogen deposition and global warming could exacerbate the volume of GHGs emitted from thawing permafrost [48]. In this research, soil samples were procured from a peatland in the Greater Khingan Mountains, Northeast China. Using laboratory incubation experiments, we assessed the impact of nitrogen supplementation on the release of greenhouse gasses and the underlying microbial processes. We propose the following hypotheses: (1) Nitrogen addition will stimulate the emission of CO2 by accelerating the decomposition of soil organic matter. (2) Nitrogen addition will enhance the abundance of methane-producing bacteria by providing more nitrogen substrates, thereby facilitating the production of CH4. (3) Nitrogen addition will promote the emission of N2O by intensifying the rate at which nitrification and denitrification processes are performed. The primary aim of this study was to explore the variations in the emission of CO2, CH4, and N2O from permafrost peatlands, as well as reveal the potential microbial processes driving these changes. This study provides a valuable reference for future predictions of soil carbon and nitrogen cycling responses to atmospheric nitrogen deposition in permafrost regions.
4. Discussion
The greenhouse gas emission rates varied greatly in the early and middle stages, with lower values in the late stages of incubation. After each gas collection, the incubation bottle would be opened temporarily to allow the soil to fully interact with the external air. Therefore, oxygen was not a limiting factor for soil mineralization [66]. Due to the complex mechanisms of soil microorganisms and the changes in soil substrates in different incubation stages, the addition of nitrogen and water had a strong effect on the soil’s microbial activity in the early and middle incubation stages, and the labile organic matter in the soil provided sufficient substrates for soil microorganisms [67]. Towards the end of the incubation period, it is likely that the reduction in the greenhouse gas emission was caused by the depletion of readily available soil organic carbon pools [68].
We found that the addition of low and high levels of nitrogen significantly increased the cumulative emission of CO2 (Figure 2b). This finding was similar to results obtained in situ on the Sanjiang Plain of Northeast China and in a Moso bamboo forest [69,70]. Interestingly however, there was no significant difference in soil CO2 emissions when comparing the control treatment with the medium nitrogen addition treatment, despite increased soil CO2 emissions by only 2.35% in the latter treatment, which was lower than the emissions from the low nitrogen addition treatment. This may be explained by the relatively high moisture content throughout the incubation. Within certain limits, increased soil moisture content can stimulate microbial activity, thereby increasing CO2 emissions. However, when the moisture exceeds a certain threshold, it can lead to increased soil particle agglomeration, limiting microbial activity and consequently suppressing CO2 emissions [71,72]. Field-based experimental research indicates that the application of nitrogen can stimulate the proliferation and metabolic activity of soil microbes, consequently enhancing the release of CO2 from the soil [73]. The high abundance of bacterial functional genes can stimulate soil respiration [19]. Fungi play a crucial role in catalyzing essential processes during the breakdown of organic matter and participate in the nutrient cycling of nearly all elements necessary for plant growth [43]. The addition of nitrogen can reduce the soil pH, which is conducive to the growth of some bacterial species [74]. In this research, the abundance of soil bacterial and fungal functional genes largely increased with the addition of nitrogen. The addition of high levels of nitrogen significantly increased the abundance of soil bacterial functional genes, with a lower increase in fungi genes when high levels of nitrogen were added. This can be explained by the fact that, when a high level of nitrogen was added, the nitrogen levels may have exceeded the nitrogen demand of fungi and adversely affected the microbial activity [74].
This study found that the cumulative CO2 emissions significantly increased by 45.50% under 0.01 g kg−1 of nitrogen addition and by 90.42% under 0.04 g kg−1 of nitrogen addition, respectively. The cumulative CO2 emissions decreased by 3.4% under 0.1 g kg−1 of nitrogen addition and by 7.4% under 0.25 g kg−1 of nitrogen addition, respectively, in subtropical forests [65]. Although the total CO2 emissions varied between the two aforementioned incubation studies, there was a consistent trend: the ratio of fungal to bacterial biomass increased as the levels of nitrogen increased and when the incubation period exceeded 100 days. The ability of the addition of nitrogen to increase or decrease the accumulation of SOC mineralization could be explained by several factors, e.g., the soil water content, the organic carbon content, and the volume of N added [75]. The relatively high rates of CO2 production may also be attributed to the fact that permafrost peatland soils are more sensitive to nitrogen because microorganisms are largely limited by nitrogen during the process of organic matter decomposition in permafrost regions [76]. Consequently, the application of nitrogen has been shown to expedite the breakdown of organic matter in soil, leading to heightened CO2 emissions [77,78].
We found that the nitrogen addition treatments significantly increased the cumulative emissions of CH4 (Figure 2d). NH4+-N can inhibit the oxidation of CH4 by competing with methane monooxygenase [79,80,81]. An increase in available nitrogen can lead to elevated CH4 emissions, as it alleviates the nitrogen limitations that might have been restricting microbial activity [10]. NH4+-N can impede the process of methane oxidation by competing with the methane monooxygenase enzyme for substrate availability [82]. The addition of nitrogen can also inhibit the activity of methanotrophs by reducing the soil pH, resulting in a decrease in the oxidation of soil methane [83]. The overall CH4 emissions demonstrated a significant positive correlation with both soil ammonium nitrogen (NH4+-N) and the mcrA gene, which is associated with methanogenesis. Conversely, they showed a significant inverse relationship with the soil pH, thereby substantiating the preceding observations (Figure 6). Our results showed that the addition of nitrogen reduces the methane oxidation capacity of soil and promotes the emission of soil CH4 [29,84].
The emissions of soil N2O accumulated progressively in response to the increasing nitrogen gradient (Figure 2f). This pattern is in agreement with other ecosystems [68,85,86,87]. An increase in available nitrogen provides more substrates for nitrifying and denitrifying microorganisms (nirS- and nirK-type denitrifying bacterial communities), promoting nitrification and denitrification and increasing the emission of soil N2O [32,88]. In our research, the total N2O emissions exhibited a significant positive correlation with the levels of nirS, nirK, and NH4+-N in the soil, while an inverse relationship was observed with the soil pH. These results confirm that a lower soil pH can significantly increase the production of N2O through enhanced denitrification; this is because a low soil pH increases the number and activity of soil-denitrifying bacteria [89,90].
This study analyzed the effects of nitrogen addition at varying levels on greenhouse gas emissions and microbial mechanisms through indoor incubation, yet with some limitations. Specifically, as the incubation period was only 154 days, it could not fully represent field conditions in terms of temperature and moisture variations nor potential interactions among microbial species. Therefore, in order to simulate field conditions more accurately, experimental setups with diverse environmental conditions (e.g., temperature and humidity variations) should be established in outdoor settings, such as open-top chambers (OTCs), to observe greenhouse gas emission patterns. Additionally, high-throughput sequencing technology can be used to investigate the composition and functional diversity of soil microbial communities. Analysis of microbial interactions through this approach can reveal the specific paths of different microbes influencing greenhouse gas emissions [91].
Source link
Boquan Lu www.mdpi.com

