1. Introduction
Endocrine-disrupting chemicals (EDCs) are substances of either natural or artificial origin that mimic or modify hormonal actions. EDCs are found in numerous widely used products including plastics, detergents, building materials, cosmetics, and pesticides. Organisms can be exposed to these hazards via inhalation and feeding, through the skin, the placenta, and the maternal milk during lactation [
1]. Due to their rudimentary detoxification mechanisms, embryos and developing organisms are more vulnerable to the effects of these hormone-mimicking chemicals. Accumulating evidence shows that EDC exposures during critical time windows can modify the developmental process of certain systems, leading to long-lasting and even transgenerational aberrations [
2].
The mandible or lower jaw differs from other bones in terms of biomechanics, morphology, physiology and morphogenesis. The mandible is the only mobile bone of the skull and has an important role in supporting dentition and in the outfit of the temporo-mandibular joint. Mandible formation is an early developmental process taking place between gestational weeks 7 and 14 in humans [
3] and embryonic days E13.5 and 16.5 in mice [
4]. During craniofacial development, neuroectoderm-derived neural crest cells interact with cells of ectoderm and paraxial mesoderm and migrate towards maxillary and mandibular prominences where they contribute to the formation of Meckel’s cartilage. The latter provides the template for mandibles, teeth, and other craniofacial structures [
5]. These early cellular interactions require tight synchronization between cell proliferation and differentiation to avoid jaw aberrations that may permanently impact its shape and function. The process of oral bone formation and odontogenesis is sensitive to external interventions including chemicals and hormones. Rodents exposed in utero to ethanol [
6], nicotine [
7], or thyroxin [
8] exhibit impaired mandible morphology at later life stages. Estrogens, apart from being significant contributors to the formation of the appendicular skeleton [
9], participate in craniofacial bone development [
10,
11]. Evidence from estrogen receptor alpha (ERα) knockout mice (ERαKO C57BL/6) shows that estrogens, via ERα, mediate the maturation of mandibular condylar cartilage in males [
12].
Consequently, estrogen-mimicking EDCs have been investigated for their effects on bone development. Perinatal exposure to Bisphenol A (BPA) or phthalates leads to impaired non-facial bone development in several species [
13,
14]. Regarding mandibular development, BPA and phthalates have been reported to affect the facial [
15] and non-facial skeletal growth [
16] in zebrafish. In rats, perinatal exposure to BPA disturbed amelogenesis in male offspring by altering estrogenic signaling [
17,
18]. Exposure of adult mice to di-(2-ethylhexyl) phthalate (DEHP) caused enamel defects in their continuously growing incisors [
19]. In humans, the developmental impact of phenols and phthalates has been examined on total body (less head) bones, where maternal levels of these chemicals during pregnancy have been associated with decreased bone mineral density in their progeny [
20,
21,
22].
Humans are exposed to chemical mixtures rather than single compounds [
23]. In previous studies, we have composed human-relevant mixtures based on epidemiological data from the Swedish Environmental, Longitudinal, Mother and Child, Asthma and Allergy (SELMA) pregnancy cohort study [
24]. Using weighted quantile sum (WQS) regression, we identified EDC mixtures measured in blood or urine of SELMA women around pregnancy week 10 associated with health outcomes in their children. Further analyses of these mixtures in different cell and animal models revealed significant effects on reproduction [
25], metabolic growth [
26,
27,
28] and neurodevelopment [
28,
29,
30]. We have previously reported neurodevelopmental effects of Mixture N1, an EDC mixture associated with language delay at 30 months, by identifying that in utero exposure to Mixture N1 altered the brain transcriptome and epigenome and, consequently, the behavior of mice offspring in adulthood [
29,
30].
Considering the impact of Mixture N1 on neural tissue development, we aimed to examine whether this mixture has an effect on the development of the mouse mandible, a structure originating from cranial neural crest cells. For this reason, pregnant mice were exposed daily throughout gestation to Mixture N1 at doses of 0.5×, 10×, 100× and 500× (representing times of human serum concentrations measured or estimated in pregnant women of the SELMA cohort study) or the vehicle. We then performed micro-Computed Tomography (micro-CT) based analysis in the mandibles of adult male offspring to accurately quantify potential changes in mandibular morphometry and bone composition. Histological staining was applied to reveal the areas of mature hypertrophic chondrocytes in the condylar head.
3. Discussion
Mixture N1 composition (DEP, DBP, DBzP, DIDP/DPHP) BPA, TCP, 3-PBA and p,p′-DDE), was defined by associating maternal levels of exposure to chemicals during pregnancy with the neurodevelopmental progress of their children in the SELMA study [
24]. We have previously demonstrated the significant impact of Mixture N1 on neurodevelopment upon gestational exposure in mice [
29,
30]. The present study showed a developmental effect of Mixture N1 on structural parameters of the murine mandible, which exhibits a cranial neural crest origin. Specifically, in utero N1 exposure modified the distances between several landmarks of the mandible and the crown width of the M1 molar, it reduced the trabecular alveolar bone volume (BV), whereas it increased the BV in the cancellous bone and altered the composition of the chondrocyte zone of the condylar head.
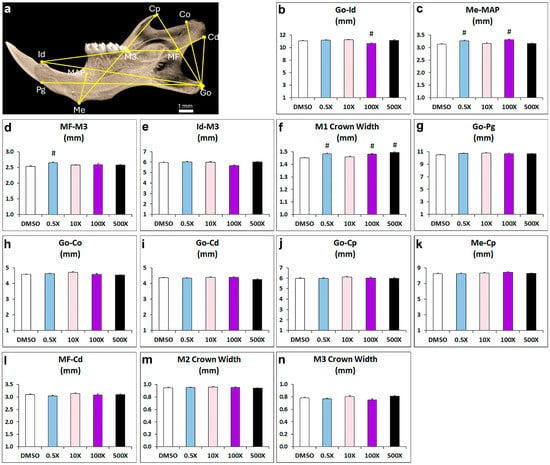
Mandibular shape and size are critical for the proper functioning of the temporomandibular junction and mastication. Consequently, changes in shape and size may lead to malocclusion. Our results showed altered distances between cephalometric landmarks of mandibular length (gonion to infradental, infradental to M3 molar and mandibular foramen to M3) and width (menton to MAP) in the Mixture-N1-exposed mice. This suggests a potential deformity of mandibular shape that could have influenced the increase in the width of the M1 molar, also witnessed in several of the experimental groups. Many of the detected effects exhibited a non-monotonic pattern that has often been reported to characterize the action of EDCs [
31,
32].
Mandibular composition provides an ideal microenvironment to support dentition and at the same time to endure multidirectional forces during chewing. Accordingly, the ramus and the angular process consist of cortical bone, while the alveolar region that hosts the teeth, and the condylar process consists of trabecular bone. In our study, the impact of Mixture N1 was detected in the trabecular areas of the alveolar bone and the condylar head. The reduced alveolar bone volume in teeth sockets contributes to periodontium loosening and potential teeth loss. Although studies linking phthalates with periodontitis are scarce, a recent epidemiological study associated urinary phthalate metabolites with increased odds of periodontitis in adult subjects [
33]. The differential impact of Mixture N1 on the alveolar bone volume fraction (reduced) and the cancellous bone of the condyle (increased) may reflect the different ossification modes of these structures. The alveolar bone at the intermediate part of the mandible (in between the molar roots) is formed by intramembranous ossification, while the condyle follows endochondral ossification [
34]. The condylar cartilage serves as a site of condylar growth through the hypertrophic chondrocytes that transdifferentiate into osteoblasts in the deeper layers of the condyle [
35].
Our results showed a reduced area of hypertrophic chondrocytes in the condyle of groups exposed to 0.5×, 10× and 100× Mixture N1. It has been shown that condylar cartilage growth in both sexes of mice is supported by gonadal steroids, especially by estrogens via ERa [
12,
36]. It is also known that components of Mixture N1 such as BPA and phthalates interfere with estrogen signaling [
37,
38]. Although direct comparisons cannot be made between the effects of single EDCs and EDC mixtures, evidence from mice perinatally exposed to BPA (200 μg BPA/kg BW) shows reduced bone volume in the trabecular femoral bones of adult male offspring [
39]. In our study, prenatal exposure to a lower dose of BPA (74.99 μg/kg/kg BW in the 500× group of Mixture N1 [
26]) had similar effects in the mandibular alveolar bone. Furthermore, BPA could have also acted as an androgen receptor (AR) antagonist [
40], counteracting the protective effects of androgens on trabecular bone mass exerted via AR signaling in osteoblasts [
36]. Nevertheless, the effects of EDC mixtures cannot be directly extrapolated to those of single compounds due to the potential mutual interactions among the contributing chemicals [
41]. These interactions could account for the lack of effects on cortical bone in our study, although it has been shown that developmental BPA exposure increases the cortical thickness of femoral bones in adult male rat offspring [
42] that exhibit the same ossification pattern with mandibular cortical bone.
Strengths and Limitations
To our knowledge, this is the first study to report impaired development of mandibular components in a mammalian model upon prenatal exposure to a human-relevant EDC mixture. Further studies are required to delineate the mechanisms of in utero effects of EDC mixtures in oral osteogenesis and dentition, since developmental alterations in mandible/condyle growth may lead to inadequate morphology and function in later life, leading to malocclusion and other complications [
43]. Given that mandibular alterations were detected in mice offspring even at low doses corresponding to the 0.5× and 10× (times) of the exposure concentrations measured in pregnant women of the SELMA cohort study, it would be relevant to include dental examinations in the follow-up of children from such cohorts.
5. Methodology
5.1. Mixture N1
A detailed description of the establishment of Mixture N1 can be found in our previous publications [
29,
30]. In brief, the urine and serum levels of 26 suspected or known EDCs or their metabolites detected in 2354 women of the SELMA study at median pregnancy week 10 were subjected to weighted quantile sum (WQS) regression analysis to select those chemicals that were associated with language delay in children at 2.5 years of age. The daily intake (DI) of the selected chemicals, the plasma concentrations from the DI estimates, and the geometric means of both urinary and serum of these chemicals were then used to determine their mixing proportions. The composition of Mixture N1 is shown in
Table 1. Phthalates’ active monoesters were used to prepare the mixture for mice. The chemicals were purchased from the following sources: Bisphenol A (BPA; 99%), Dimethylsulfoxide (DMSO; 99.9%), Monobenzyl phthalate (MBzP; 98%), 3-Phenoxybenzoic acid (3-PBA; 98%), and Trichloropyridinol (TCP; 99%)were obtained from Sigma-Aldrich Inc. (St. Louis, MO, USA). Monoethyl phthalate (MEP; 98%)and mono-iso-decyl phthalate (MiDP; 98%) were obtained from Toronto Research Chemicals (North York, ON, Canada).Mono-(2-propylheptyl)-phthalate(MPHP, 99%) were synthesized by Novandi Chemistry AB, Södertälje, Sweden. Monobutyl phthalate (MBP; 95%) was purchased from TCI, Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan). pp′DDE was synthesized in house by Åke Bergman from DDT. For MIX N1, 1 M solutions in DMSO were prepared of each of the chemicals: BPA, MEP, MBP, BBzP, MIDP, MPHP, 3-PBA, and TCP. A 50/50 mixture of MIDP and MPHP was used [
29].
5.2. Experimental Protocol and Tissue Preparation
In the present study, we examined the effect of prenatal exposure to Mixture N1 on the mandibles of three-month-old male offspring. The animal tissues and data were collected in a previous study for Mixture N1 [
29]. The study’s protocol was approved by the Ethical Committee of the Prefecture of Attica-Veterinary Department (#4783) and performed in accordance with the European Communities Council Directive of 22 September 2010 (2010/63/EU). In brief, two-month-old C57/BL6 mice, purchased from the Hellenic Pasteur Institute (Athens, Greece), were used for breeding. The animals were housed under standard conditions of temperature and illumination and were offered a phytoestrogen-deficient pellet food (Altromin 1324P, Lage, Germany) and tap water ad libitum. From gestational day 1 to parturition, pregnant mice received daily via food the vehicle (Dimethylsulfoxide, DMSO) or the N1 mixture at doses of 0.5×, 10×, 100× and 500× hsc (
times of human serum concentration represents the exposure concentrations relative to the geometric mean of the concentrations measured in pregnant women of the SELMA cohort study). Accordingly, the daily exposure of pregnant dams throughout pregnancy was 0.001, 0.22, 2.2 and 11 mg/kg bw of Mixture N1, respectively. Working solutions of different doses were prepared using DMSO 99.9% purity, Sigma-Aldrich Inc. (St. Louis, MO, USA). For all pregnant mice, DMSO intake did not exceed 0.25 μL/gr bw/day [
29], which is considered a non-toxic exposure. Offspring were euthanized at three months of age under isoflurane anesthesia. The mandibles were removed and fixed in neutral formalin. One hemimandible was dehydrated and kept for micro-CT analysis. The respective condyles from the other half were decalcified in 10% EDTA solution, washed in tap water, dehydrated, and embedded in paraffin. The number of samples per group used in the analysis was: DMSO: n = 10, 0.5×: n = 8, 10×: n = 10, 100×: n = 7, 500×: n = 11.
5.3. Micro-CT Analysis
The microarchitecture of the mandibular bones was evaluated using a high-resolution SkyScan1172 microcomputed tomography (mCT) imaging system (Bruker, Aartselaar, Belgium). Images were acquired at 60 KeV, 100 µA, pixel size 10 μm, with a 0.5 mm aluminum filter. Two-dimensional reconstruction images were generated using Nrecon software (version 1.7.4.6) (Bruker) and analyzed using Ctan software (version 1.20.8) while 3D reconstructed images of alveolar bone were generated using Ctvox software (version 3.3.1) (Bruker). Morphometric measurements were performed with Data Viewer software (version 1.6.0) (Bruker micro-CT). Analyses were performed between the following reference points: gonion (Go) to dorsal point of the condylar process (Co); Go to ventral point of the condylar process (Cd); Go to coronoid process (Cp); Go to the most antero-dorsal point on mandibular symphysis (pogonion, Pg); Go to the most postero-dorsal point on mandibular symphysis (infradental, Id); menton (Me) to mandibular alveolar point (MAP); Me to Cp, mandibular foramen (MF) to Co; MF to M3 molar; Id to the most posterior part of M3 molar (
Figure 1a). The crown width of M1, M2 and M3 molars was also measured.
Alveolar, trabecular, and cortical bones were analyzed from transaxial sections, respectively, between the roots of the first molar, at the mandibular condyle, and at the posterior edge of the ramus. The parameters of bone volume (BV, mm3), tissue volume (TV, mm3), bone volume fraction (BV/TV, %), trabecular number (Tb.N, mm−1), trabecular separation (Tb.S, mm), trabecular thickness (Tb.Th, mm) and bone mineral density (BMD, g/cm3) were measured for alveolar trabecular bone and condyle head. Cortical bone was assessed through cortical bone volume (Ct.BV, mm3), tissue volume (TV, mm3), cortical thickness (Ct.Th, mm), and tissue mineral density (TMD, g/cm3).
5.4. Histomorphometry of Condylar Head
The basic histology of the condylar head was examined in 4 μm paraffin sections (Leica RM 2125 microtome, Leica Microsystems, Wetzlar, Germany) stained with Hematoxylin and Eosin (H&E). Safranin-O/Methylene green staining was used to locate the zone of hypertrophic chondrocytes in the condylar head. Digital images of stained sections, corresponding to the mid-coronal part of the condylar head, were obtained by optical Microscope (Eclipse E400, Nikon, Tokyo, Japan) and the stained areas were quantified as % of the total condylar head area using ImageJ Software (v. 1.54f, 2023, NIH Image, National Institutes of Health, Bethesda, MD, USA) by two independent observers blindly. Animal samples used in quantification of staining: DMSO: n = 6; 0.5×: n = 5; 10×: n = 4; 100×: n = 6; 500×: n = 4. For each sample, an average of 3–5 sections was used in the analysis. In addition, the area width of hypertrophic chondrocytes was measured in the same samples based on the morphology of these cells. Specifically, for each available section from each animal/group, we performed 8–12 (usually 10) vertical measurements in the hypertrophic chondrocytes’ area, in most cases following the palisades formed by the hypertrophic chondrocytes. We used a systematic-random sampling method starting at a random point at the left end of the zone and measuring every 50 μm (lateral step), to cover the whole length of the zone.
5.5. Statistical Evaluation
Data were analyzed by generalized linear models (GLMs), with the dose of Mixture N1 (treatment) as the predictor factor and the treatment (litter) as a build nested predictor factor (IBM Statistical Package for the Social Sciences (SPSS) v. 21). In the case of statistically significant dose effects, Bonferroni post hoc tests were used to determine specific group differences compared to the DMSO-treated group. To control for type I error due to multiple statistical comparisons, an adjusted p-value threshold (p < 0.0015) has been calculated dividing p = 0.05 by the total number of statistical comparisons performed.