1. Introduction
Maize (
Zea mays L.) is a vital crop cultivated worldwide under diverse climatic conditions [
1], serving as a staple food for millions, as well as a key resource in animal feed and various industrial applications [
2]. However, maize growth is often impeded by several environmental factors, such as salt stress, extreme temperatures (both high and low), toxic metal exposure, UV radiation, and drought stress [
3], all of which adversely affect plant growth and productivity depending on the severity of the stress. In response to these stresses, plants have developed a range of physiological and biochemical mechanisms to increase their tolerance and survival.
Among these stresses, water deficit, or drought stress, is a major abiotic factor that severely limits plant growth and yield. Drought stress induces the production of reactive oxygen species (ROS), including hydroxyl radicals (‧OH), superoxide anions (O
2¯), and hydrogen peroxide (H
2O
2). The overaccumulation of ROS leads to oxidative stress, causing significant damage through membrane lipid peroxidation [
4], weakening the plant’s defense system, and resulting in protein oxidation, nucleic acid damage, enzyme inhibition, and, ultimately, programmed cell death [
5]. Drought stress is particularly detrimental to crops, as it decreases their quality, yield, and stability, especially when experienced during critical stages of growth [
6]. In rice and other crops, drought negatively impacts the initiation of flowering, leading to panicle sterility and reduced yields [
7]. With the increasing global demand for food, developing drought-tolerant varieties has become crucial. Understanding the biochemical and physiological mechanisms that enable plants to withstand drought stress is essential for selecting and breeding crops with enhanced drought resilience [
8]. Under drought stress, plants employ various strategies to cope, including evading drought conditions and enhancing tolerance through specific adaptations. Osmotic adjustment, in particular, is a key physiological process critical for maintaining plant growth under drought stress [
9].
Several studies, as summarized by Qasim Ali et al., have reported that plants accumulate various osmoprotectants, such as proline, to enhance their tolerance to both salt and drought stress [
8]. Proline functions as a key osmoprotectant, stabilizing protein structures and the photosynthetic apparatus [
10]. It also aids in regulating cellular osmotic balance and mitigating the effects of ROS during abiotic stress conditions [
11,
12,
13,
14]. Proline scavenges ROS, including H
2O
2 and O
2¯, generated during drought stress, thereby alleviating oxidative stress and protecting the plant from associated damage [
15]. Additionally, proline enhances turgor potential, photosynthetic activity, and antioxidant activity, resulting in reduced oxidative damage [
16]. Yang et al. [
17] and Ihtisham et al. [
18] have shown that proline can effectively mitigate the detrimental effects of oxidative stress in plants. Plants that accumulate higher levels of proline demonstrate enhanced stress tolerance [
19,
20], promoting growth by reducing nutrient imbalances, lowering elemental toxicity, and enhancing photosynthesis [
21]. Exogenous proline application during the seedling and vegetative stages of maize has been shown to promote growth under drought conditions [
8]. Furthermore, pre-sowing seed soaking with proline has been reported to enhance growth in
Triticum aestivum under drought stress [
22]. Proline supplementation also helps to maintain optimal nutrient (potassium, calcium, phosphorus, and nitrogen) levels in maize, contributing to improved drought tolerance [
23]. Potassium is particularly vital for key physiological processes in plants, including stomatal function, transpiration, photosynthesis, photophosphorylation, turgor pressure maintenance, enzyme activation, and photoassimilate transport [
24]. Recent studies indicate that applying potassium externally can mitigate the adverse effects of drought on rice, supporting key physiological functions, including protein synthesis, enzyme activity, photosynthesis, water regulation, and stomatal control [
24].
Although proline is known to play a significant role in plant growth and development under various environmental stresses, its specific impact on maize plants during drought stress remains underexplored. We hypothesized that proline supplementation could alleviate the detrimental effects of drought stress on maize by enhancing its resilience. The primary objective of this study was to investigate the role of proline in modulating key morpho-physiological and biochemical parameters in maize exposed to drought conditions. Specifically, we aimed to assess whether proline could improve growth, antioxidant enzymatic activity, water retention, nutrient balance, and photosynthetic efficiency, thereby promoting overall plant performance and stress tolerance. The results of this study provide valuable insights into the use of proline as a strategy for enhancing drought resistance in maize.
4. Discussion
Drought stress significantly limits maize productivity by impairing various physiological, biochemical, and molecular processes. Our study demonstrates that proline supplementation enhances maize resilience to drought by improving growth, water retention, membrane stability, and nutrient accumulation while reducing oxidative damage (
Figure 6 and
Figure S1). These findings highlight proline’s multifaceted role in mitigating drought stress and offer valuable insights for developing strategies to improve crop resilience under challenging climate conditions.
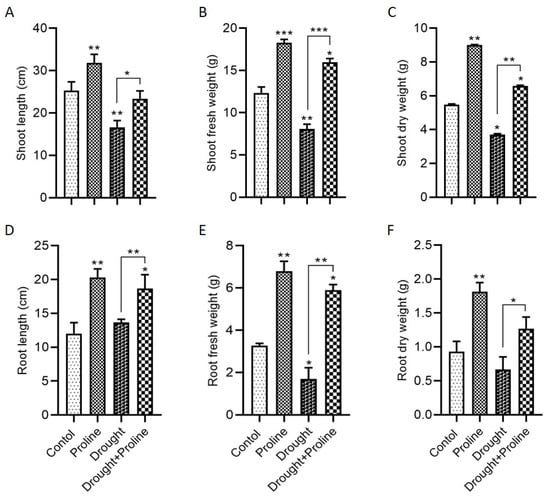
Plants employ several adaptive mechanisms to cope with environmental stresses, particularly abiotic stresses like drought. The application of osmoprotectants, such as amino acids, proline, betaines, and trehalose, strengthens plant resilience by reducing stress-related damage [
36]. In this study, we investigated the effect of exogenous proline on maize plants under drought conditions, demonstrating its role as an efficient osmolyte. Proline supplementation significantly improved plant growth parameters, including shoot and root lengths, fresh and dry weights, leaf area, and stem diameter, under drought stress. Importantly, exogenous proline application increased endogenous proline accumulation in both the roots and shoots, further supporting growth under drought conditions. In contrast, drought stress alone severely impaired these growth parameters. These results align with previous studies showing that exogenous proline application in maize promotes endogenous proline accumulation and enhances growth, mitigating the detrimental effects of drought stress [
8]. This is consistent with findings in rice and
Allenrolfea occidentalis, where proline supplementation enhanced growth under both drought and salt stress [
37,
38]. Exogenously applied proline boosts endogenous proline levels, stabilizing enzymes, proteins, and membrane components, while potentially providing energy for growth and stress tolerance during drought [
8]. Similar to drought stress, exogenous proline supplementation has been shown to enhance plant growth and biomass under salt stress in various species, such as
Medicago sativa,
Helianthus annuns,
Cucumis sativus,
Triticum durum,
Zea mays, and
Oryza sativa [
39,
40,
41,
42,
43,
44]. However, the impact of exogenous proline can vary depending on the plant species, developmental stage, application method, and concentration. In our study, applying 30 mM proline at the seedling stage significantly improved maize growth parameters, mirroring findings in rice, where 30 mM proline enhanced growth at the seedling stage. Comparable results have been reported with 30 mM proline in mung bean cell cultures, while 10 mM was effective in tobacco suspension cells [
45,
46]. These findings highlight the potential of proline as a versatile osmoprotectant that enhances plant growth and resilience under drought conditions, especially when applied at optimal concentrations and developmental stages. Further research could explore the underlying mechanisms of proline-induced stress tolerance, offering valuable insights for developing sustainable crop management practices in stress-prone environments.
In maize, drought stress markedly affects photosynthesis by reducing leaf area and chlorophyll levels, both of which are crucial for maintaining high photosynthetic efficiency. In our study, limited water availability led to a decline in chlorophyll levels, primarily due to increased oxidative stress, resulting in elevated H
2O
2 and MDA levels, as well as reduced activity of antioxidant enzymes, including CAT, POD, and SOD (
Figure 3). However, exogenous application of proline effectively alleviated oxidative stress, enhancing antioxidant enzyme activities and thereby mitigating drought-induced damage. Additionally, proline anabolism can enable plants to maintain osmotic balance, facilitating the recovery of water content, especially during osmotic stress [
47]. Similarly, Qasim Ali et al. demonstrated that proline application supports photosynthesis in maize under drought conditions by mitigating oxidative stress [
8]. Our findings further suggest that proline application reduces H
2O
2 production and MDA accumulation (
Figure 3A,B), which in turn enhances stomatal conductance and CO
2 assimilation, thereby promoting photosynthesis. These observations are in line with previous studies showing that foliar application of proline in drought-stressed maize improves stomatal conductance and CO
2 uptake, supporting photosynthetic processes [
48].
Additionally, our study indicates that exogenous proline increases cellular proline levels, which stimulates the activities of antioxidant enzymes, such as CAT, POD, and SOD, effectively reducing oxidative stress. Several studies have suggested that proline functions as an electron acceptor, boosting antioxidant enzyme activity and minimizing oxidative damage [
49,
50,
51]. For example, in sugar beet, exogenous proline has been shown to increase the levels of endogenous proline and phenolic compounds, as well as antioxidant enzyme activities, helping to alleviate oxidative stress symptoms [
52]. Furthermore, numerous studies have indicated that proline application can boost stress tolerance by improving nutrient acquisition, water uptake, photosynthesis, gas exchange, and antioxidant responses [
47,
53,
54,
55]. In our study, proline application under drought conditions mitigated the buildup of H
2O
2 and MDA by enhancing antioxidant enzyme activities. Similar results were observed in recent studies, where seed priming with proline enhanced ascorbate peroxidase (APX), CAT, and SOD activities in maize cultivars FS-67 and CML-539 [
56]. Notably, CAT triggers various responses that help plants adapt to stress, while SOD acts as a frontline defense by quickly converting ROS into safer molecules, such as oxygen and water [
57]. A recent study also indicated that proline acts as an ROS scavenger, protecting chloroplasts and nuclei from oxidative damage [
58]. These results highlight the potential of proline as a valuable osmoprotectant for alleviating drought-induced oxidative stress, enhancing photosynthetic efficiency, and improving overall plant resilience under water-deficit conditions.
Under drought stress, plants employ various adaptive mechanisms to maintain cell turgor, one of which is osmotic regulation. This process involves the accumulation of osmoprotectants, such as proline, soluble sugars, and amino acids, to help retain water and preserve cellular function. Studies have shown that osmolytes play a vital role in plant defense by enabling cells to retain water via osmotic adjustment, thereby preserving turgor pressure [
59,
60]. In our study, maize seedlings treated with exogenous proline under drought conditions exhibited significantly higher levels of total protein, soluble sugars, and essential nutrients (N, P, and K), along with increased intercellular proline, compared to the untreated drought-stressed plants (
Figure 4 and
Figure 5). Although some studies suggest that drought stress alone can elevate the levels of proteins, sugars, and amino acids compared to those observed in irrigated plants [
55], our findings revealed a reduction in these metabolites in plants subjected solely to drought stress compared to the control plants. This discrepancy may be attributed to the higher severity or prolonged duration of stress in our study, potentially overwhelming the plants’ adaptive mechanisms. Differences in plant responses to drought stress across studies are likely influenced by factors such as plant species, developmental stage, environmental conditions, and experimental designs, all of which shape metabolic adaptations to stress. In the present study, proline-primed maize seeds accumulated elevated levels of endogenous proline and sugars during drought conditions [
16], consistent with other studies indicating that exogenous proline application increases endogenous proline levels in water-stressed plant tissues, enhancing osmotic adjustment [
8,
36,
61,
62].
Our results also align with recent studies demonstrating that exogenous proline promotes the uptake of potassium, calcium, phosphorus, and nitrogen under drought stress [
23], highlighting the importance of nutrient balance for drought tolerance. Potassium is essential for osmoregulation, enzyme activation, and stomatal regulation, while nitrogen contributes to cellular structures, and phosphorus plays a central role in energy transfer, respiration, and photosynthesis [
63,
64]. Notably, in our study, drought stress without proline supplementation significantly reduced the levels of these key nutrients (N, P, and K). Pessarakli [
65] attributed such reductions to lower respiration rates and stomatal conductance during water stress. Given that nutrient uptake in plants depends on water movement from the roots to the shoots [
66], our findings suggest that exogenous proline application mitigated drought stress in maize by promoting nutrient absorption and accumulation. This suggests that proline application may serve as an effective strategy for improving drought tolerance in maize by improving the levels of essential nutrients under water-limited conditions.