1. Introduction
The Arecaceae family (palms), with around 2400 species, is widely distributed in tropical and subtropical regions [
1,
2,
3]. In Mexico, around 95 species of palms have been recorded [
4], 20 of which are distributed in the Yucatan Peninsula [
5,
6]. Given their significant economic and socio-cultural importance in this area, many studies focus on palm production, uses, and management [
7,
8,
9,
10,
11,
12,
13]. However, information regarding the conservation status (and necessary restoration actions) of this group of plants is generally lacking, and in fact, several palm species are already in some endangered category due to over-extraction, disturbance, and habitat loss [
14,
15]. In this sense, ecological studies of this group of species are limited, and most of them aim at understanding the effect of disturbances in populations and knowing the factors that limit the distribution of some species [
1,
5,
6,
14]. There are no studies aimed at understanding the impact of environmental variables (such as type of substrate) on palm groves, which would be helpful for management and conservation programs.
In Mexico, most vegetation (floristic) studies that include palm species focus on the continental region of the Yucatan Peninsula, and very few include the vegetation of the Mexican islands. For example, some authors only mention the presence of
Thrinax radiata (hereafter referred to as
Thrinax) on the Island of Cozumel in the Mexican Caribbean [
16,
17,
18,
19,
20,
21]. In particular, the palm grove community of
Thrinax has only been superficially studied in the interior part of the island [
17]. However, its structure and species composition are largely unknown, especially for the palm groves growing on the coastal area in the eastern part of the island, where the palm is more frequent and abundant. The presence of these communities stabilizes the dunes and accumulates organic matter that promotes soil formation. Furthermore, their structure and dense cover reduce wind- and wave-induced erosion and help protect inland vegetation and human infrastructure [
18].
Usually, palm grove communities are not very diverse, and one palm species is often dominant [
6,
22,
23]. Several environmental characteristics determine the distribution and abundance of different palm species, such as environmental relative humidity, soil moisture, and soil type, among other factors [
2]. The palm grove of Cozumel is distributed along the eastern coast in the protected natural area of the island and discontinuously south along the transversal road until the area known as Punta Sur [
18,
19]. The sandy substrate is dominant in this eastern coastal area, although gley soils are also present [
24]. These soils have little organic matter and great permeability, but the depth of the sandy substrate is greater than the rocky gley soil in which outcrops of limestone sedimentary rock emerge [
24].
Based on the above, the study goals of this study were (1) to describe the palm grove communities present on the eastern coast of the island of Cozumel and (2) to explore the role of different soil types in shaping community structure and composition of the Thrinax palm grove. We hypothesized that (a) palms would be dominant in the palm grove, independent of the soil type, and (b) because the rocky gley soil is shallower than the sandy substrate and thus limits plant growth, we expected a reduced species richness and lower palm cover and density in the rocky site than on the sandy one. To our knowledge, there are no studies exploring soil influence on palm groves of Thrinax radiata.
2. Results
2.1. Community Composition
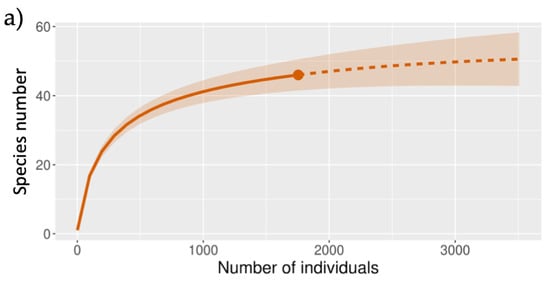
We found 27 families, of which 46 species were woody, including 24 trees, 19 shrubs, and 3 vines, with a total surface of 2900 m
2 (
Table 1). The rarefaction curve indicates that the sampling effort was sufficient since the sample coverage was 0.995 (
Figure 1a). The values of Chao 2 and Jackknife 1 suggest the potential presence of a maximum of 54 species for this plant community.
The hierarchical clustering analysis generated two groups that separated the sampling plots from each type of substrate according to their floristic composition (
Figure 2). The first group comprised 17 sampling plots, almost all corresponding to the sandy substrate, while the second group comprised 12 sampling plots, almost all from the rocky substrate. These groups were used in the following analyses and comparisons. The rarefaction curves indicated that the sampling effort was more complete in the rocky substrate than in the sandy substrate (
Figure 1b).
As expected,
Thrinax radiata was the most abundant species in both substrates. The two palm groves differed in 33% of their species, most of which were represented by a small number of individuals (
Table 1). The two floristic groups shared 20 species, representing 67% of the community. Some species, such as
Cordia sebestena,
Gymnanthes lucida,
Randia aculeata, and
Sideroxylon americanum, were more abundant in the sandy substrate, whereas
Neea psychotrioides and
Pscidia piscipula were more abundant in the rocky substrate. Hill numbers at 95% sampling efficiency indicated differences between palm groves established in the two substrates. The species richness (
0D) and the abundant (
1D) and dominant (
2D) species numbers were higher in the rocky substrate than in the sandy substrate (
0Dsandy = 15.2,
0Drocky = 21.1;
1Dsandy = 2.5,
1Drocky = 3.7;
2Dsandy = 1.5,
2Drocky = 1.9).
2.2. Community Structure
Although the palm groves from both substrates had the same number of species (33 each), the community structure differed between them because the proportion of the different growth forms varied. The palm grove growing on the sandy substrate contained 18 species of trees, 12 shrubs, and 3 vines. In contrast, the palm grove from the rocky substrate had 18 species of trees, 15 shrubs, and no vines.
The importance value index (IVI) showed that
Thrinax radiata was the most important species in the two substrates (
Figure 3), thus defining the palmar community. However, the importance value of
Thrinax on the sandy substrate was much higher than its value on the rocky substrate (57 and 36, respectively) (
Figure 3). The subsequent species with higher IVI had notably lower values than those obtained for
Thrinax and varied between the two substrates. After
Thrinax, the most important species on the rocky substrate were
Neea psychotrioides,
Piscidia piscipula,
Trichilia americana, and
Diospyros verae-crucis, among other species. In contrast, after
Thrinax, the most important species of the palm grove growing in the sandy substrate were
Cordia sebestena,
Neea psychotrioides,
Sideroxylon americanum,
Pithecellobium keyense, and
Randia aculeata, among others. The species found in the two substrates maintained low importance values. However, some had higher values in the sandy substrate
(Cordia sebestena) and some in the rocky substrate
(Neea psychotrioides, Piscidia piscipula, and
Pithecellobium keyense). One species,
Harpalyce arborescens, had similar importance values in the two substrates (
Figure 3).
The density of plants, mean plant cover, and mean DBH per plot also varied between the substrates. We found 953 individuals on the sandy substrate and 775 on the rocky, yielding densities of 0.65 and 0.56 individuals /m
2, respectively. Most of the individuals in both substrates were
Thrinax palms (737 in the sandy substrate and 538 in the rocky substrate). Like the results for plant density, mean plant cover and DBH per plot were larger in the sandy than in the rocky substrates, although only differences in DBH were significant for all plant communities (
Figure 4a,b). These trends in DBH held for the plant community without including
Thrinax (
Figure 4c) and for
Thrinax alone (
Figure 4d).
Finally, and contrary to the findings for plant density, plant cover, and DBH, there were no significant differences in the height of the palms (mean + EE; sandy 6.3 ± 0.3 m, rocky 6.5 ± 0.4 m; t = −0.578, p = 0.568) and the mean height of the entire community without the palm (sandy 5.4 ± 0.3 m, rocky 5.0 ± 0.2 m; U = 4849, p = 0.37).
3. Discussion
Our results corresponded to the two hypotheses. Despite the community’s high species richness, we found that palms are the dominant species in the palm grove of Cozumel Island. We also proved that species richness, plant cover, and plant density were reduced in the rocky soil substrate compared with the sandy soil one. The reason for the last one is that the rocky gley soil is shallower than the sand soil, thus limiting plant growth.
3.1. Community Composition and the Dominance of Thrinax radiata
The palm grove community located on the eastern coast of the Island of Cozumel has a high richness of plant species (46 woody species), with the dominance of the palm Thrinax radiata. This is the most important and physiognomically dominant species of the plant community (in terms of numerical dominance, coverage, height, and DBH). The rest of the species are not as abundant and do not represent relevant physiognomic elements.
Palms are abundant in the Yucatan Peninsula, where 20 species have been recorded.
Thrinax is the palm with the amplest distribution in the peninsula and is conspicuous in the vegetation along with other palms [
15,
25]. However, in some places, a single species dominates the landscape [
6]. Our results are consistent with the dominance of a single palm species observed in other palm groves [
2]. However, it is striking that in the plant communities of the north and center of the eastern region of the Yucatan Peninsula, three palm species are dominant:
Thrinax radiata,
Sabal yapa, and
Chamaedorea seifrizii, which are the three most abundant palms in the peninsula [
5] growing on poorly developed soils like those existing on the island of Cozumel [
5,
6]. Thus, the absence of
Sabal yapa and
Chamaedorea seifrizii on the island may be associated with their limited dispersal by terrestrial vertebrates such as peccaries, squirrels, and mice [
26,
27,
28].
Other environmental factors may be favoring the abundance of
Thrinax and limiting the presence of other palms. For instance, tropical cyclones and hurricanes are regular meteorological phenomena in the Yucatan Peninsula; in the period of 2000–2012, 21 cyclones hit the coasts of the Yucatan Peninsula, 13 of them with a hurricane category [
29], and they frequently affected Cozumel, especially along the island’s eastern coast [
18]. Besides human infrastructure, hurricanes also have long-lasting effects on plant communities: they induce mortality and open gaps in the canopy and thus alter composition and structure [
30,
31,
32,
33,
34,
35]. Previous observations have shown that palms are tolerant to the impact of hurricanes, especially
Thrinax radiata, which had a high survival rate after Hurricane Wilma in 2005 [
36,
37]. Thus, it is likely that the dominance of
Thrinax in the palm grove of Cozumel is also associated with this species’ ability to better tolerate the limiting environmental conditions of the island compared with the other palms and associated species [
2]. The result is a
Thrinax-dominated palm grove with high species richness but a reduced abundance.
3.2. Community Structure
Our results showed that the palm groves from the eastern coast of Cozumel are very well defined by the two types of substrates (sandy and rocky) that are dominant in the area [
18,
22]. In general, these soils are poor in nutrients and have high infiltration, but even so, the accumulation of organic matter and sand, as well as the exposure of limestone rock, varies over relatively short distances. These factors are considered determinants in the richness and structure of plant communities [
38,
39] and, consequently, modify the richness of palm species in Cozumel and other continental nearby sites in the Yucatan Peninsula [
6].
Although
Thrinax radiata is the dominant physiognomic element in the two substrates and the two palm groves have the same number of species (33), there are some differences in their abundance and dominance and in the composition and structure of the two communities (
Table 2). Our results showed that shrubs are more abundant on the rocky substrate than the sandy substrate, but vines are only present in the palm grove that grows on the sandy substrate. Furthermore, plant and palm density is higher in the sandy substrate, where the tallest and thickest individuals are present, while in the rocky substrate, the plant density is lower, and plants are thinner and shorter. The low availability of soil in the rocky substrate does not allow the same vegetation development that occurs in the sandy substrate. Coastal sandy substrates can be enriched with the accumulation of leaf litter from the vegetation cover and their decomposition by crabs and microorganisms [
40,
41], and although nutrient availability could take some time [
42], a similar process may be occurring in our study area, producing the structural and compositional differences between the palm groves growing in sandy and rocky soils in Cozumel.
In particular, the reduction in the size and importance value of Thrinax radiata and the associated species growing in the rocky substrate draws our attention. Further studies are required to include the physical and chemical analyses of the soil, combined with the plant’s responses to these differences in soil characteristics. On the other hand, it is necessary to carry out focal studies to explore the unique distribution of the species that only occurred in one of the substrates (e.g., Trichilia americana in the rocky substrate and Sideroxylon americanun in the sandy substrate) to identify the factors that limit their presence in both substrates.
3.3. Caveats of the Study
This study shows how palm groves growing on rocky vs. sandy soils differ in number of shrubs, vines, plant cover, plant density, plant height, and DBH (
Table 2). However, the differences between soil types and palm groves were not clearcut. Some plots from the sandy soil were more like those from the rocky plot and vice versa. These trends are probably the result of species exchange between plots and a gradient in soil attributes. A detailed soil analysis would further help explain the differences between palm groves growing on predominantly rocky and sandy soils. For instance, besides the relative rockiness of the surface, community differences could be associated with, for example, nutrient content and depth of aerobic and anaerobic soil. In addition to soil analysis, the rates of propagule flow between plots would also provide information on the dynamics of these palm groves on a meta-community scale.
Despite these limitations, our results help distinguish how palm grove communities are heterogeneous even though one species is dominant. Such variability is most likely associated with differences in soil attributes. Experimental data would help demonstrate this correlation and find its causes.
5. Conclusions
Although
Thrinax radiata is a species of economic and cultural importance among the Mayans and is used for construction in the Yucatan Peninsula, the palm grove of Cozumel is in a good state of conservation and reaches high densities along the eastern coast of the island. The good conservation status is due to several factors. Its location in the conservation area of the island and its designation as a threatened species by the official Mexican standard [
14] have protected
Thrinax from the inhabitants’ use and over-exploitation.
Our results confirm that contrasting substrates resulted in different community compositions and structures of two palm groves, even though Thrinax was dominant in both. The palm groves from the sandy substrate had more vines but fewer shrubs and higher plant density, height, and DBH. The above suggests that the low availability of soil in the rocky substrate does not allow the same state of vegetation development that occurs in the sandy substrate.