2.1. The Faunal Dispersals as Key Bioevents for the Formation and the Evolution of the Villafranchian Faunas of Europe
The Mammuthus event. Lindsey et al. [
20] draw attention to data suggesting a similar age of the occurrence of
Equus and
Mammuthus in Eurasia. They allow the appearance of both species in Europe, in the interval 3.0–2.6 Ma, and Azzaroli [
16] summarizes this dispersal in a common dispersal event, accepted and mentioned repeatedly in the later literature. Recent data on the first appearance of these two genera in Europe show that these two events should most likely be separated because they are not synchronous (Iannucci, Sardella [
42]), although some questions in this regard remain (see below). The origin of the genus
Mammuthus is known to be African. The oldest
Mammuthus remains in Europe, with a secure dating, are known from Tuluceşti and Cernãteşti, Romania (Radulesco, Samson [
43]) and from the faunal complex at Ripa Skortselskaya, Moldova (Alekseeva [
44]) which is from an equivalent horizon (Skortselskian horizon: Nikiforova et al. [
45]), and they represent the first
Mammuthus species from Eurasia:
M. rumanus (Markov, Spassov [
46]; Lister, van Essen [
47]; Markov [
48]). The localities have been palaeomagnetically correlated to the mid-Gauss subchron (C2An2n, 3.207–3.116 Ma) (Andreescu et al. [
38]). It is also worth mentioning the mandible from Bossilkovtsi, Bulgaria (Markov, Spassov [
46]) (probably at least 3 Ma?) and the find from Tsotylio, Greece (ca. 3.2 Ma?) (Kostopoulos and Koulidou [
49]) show very primitive morphology and evolutionary stages similar to those of the mentioned finds from Romania, while at the same time, showing the Balkan route of dispersal. Thus, in Eurasia,
Mammuthus was a newcomer from Africa during the mid-Piacenzian, and its appearance in Europe is confined to 3.2 Ma (Markov [
48]; Böhme et al. [
1]).
The Equus s. lato event. Of the three taxa that have become classic examples of bioevents associated to one degree or another with the beginning of the Pleistocene,
Equus,
Mammuthus, and
Canis (Azzaroli [
16]), an example of a very rapid dispersal from the East is
Equus. Equus appeared in N. America at least at ca. 4 Ma (Rook et al. [
50]). In China, there are no known finds from earlier than 2.5 million years ago (Sun and Deng [
51]); this does not mean that the horse did not arrive there earlier, because in Europe today, there are a number of paleontological sites that indicate an earlier appearance of the monodactyl horse of the continent (the subgeneric taxonomy of
Equus, especially its early forms, and even the generic taxonomy of the monodactyl Villafranchian horses of Eurasia remains controversial; see Bernor et al. [
52] and Eisenmann [
53]). At least four European sites with the presence of primitive, stenonoid horses have an age of about 2.6 Ma (Roca-Neyra, El Rincón 1; Huélago, Montopoli); the Asia Minor site of Güliazi is also of this age, and Pardines (France) is a little younger than 2.6 Ma (Iannucci and Sardella [
42]). Here we must also add the Bulgarian locality of Varshets, where remains very close to
E. stenonis from Saint-Vallier were found (Spassov [
54]). The age of Varshets, given its fauna, placing it (Spassov [
11]) between Saint-Vallier (probably around 2.5 Ma) and Roca-Neyra (about 2.6 Ma), (see: Nomade et al. [
19]) should be at least 2.5 Ma. With this, the age of the earliest appearance of monodactyl horses in Europe and Western Asia should seem clear, but there is some, albeit debatable, evidence of an earlier age from Romanian sites. Samson [
32] described
Equus (
Allohyppus)
euxinus from Maluşteni and redescribed
E. simionecui from Bereşti. He also mentioned a “
Plesippus (
Allohippus)
athanasiui” metapodial from Capeni. This metatarsal III is noted also by Radulescu and Samson [
33,
55] as
Plesippus athanasiui and Allohippus cf.
athanasiui, respectively, and it is discussed (as
Equus cf.
athanasiui) by Forstén [
56], who presents some basic dimensions of this metapodial. In addition, Radulescu and Samson [
55] note the presence of the monodactyl horse also in Tuluceşti. Bereşti is an MN14 locality with an age of more than 4 Ma (Radulescu and Samson [
55]; Andreescu et al. [
38]), and the presence of a monodactyl horse with cabaloid features (Forstén [
57]) there seems illogical.
It is likely that the remains entered the locality deposits accidentally or were collected from the surface/near the locality. According to our observations (2023), the remains of
Equus from the locality, stored in the collection of the University of Bucharest, with an apparently cabaloid morphology, have a more different fossilization from that of the hipparion remains from the same locality. The Maluşteni locality is almost the same age, or perhaps only slightly younger, and is referred to as MN15a or the MN14/MN15 boundary (Andreescu et al. [
38]; Radulescu and Samson [
55]; Crespo et al. [
58]). At the same time, Crespo et al. [
58] noted the presence, albeit single, of
Spermophilus cf.
nogaici, which should be an indication of an MNQ16 zone. “
Equus euxinus” has primitive, stenonoid features. It is not excluded, considering
S. cf.
nogaici in the locality, that there is also a later level in Maluşteni, or most likely, separate intrusions of a later (MNQ16) fauna there. The question requires further research. The finds from the Pliocene Capeni locality (with a biochronological age referring to MN15b, according to Radulescu and Samson [
55], but possibly younger; see Forstén [
56]), and from the final Pliocene Tuluceşti locality (3.2 Ma; see above) were not found by me in the collections of the Institute of Biospeleology and the University of Bucharest during my visit in 2023, and it remains unclear where they are stored. These remains, however, should belong to monodactyl horses. It seems that the question of the first appearance of the monodactyl horses in Europe cannot yet be considered definitively resolved. It is possible that there was an initial dispersal on the continent in the Mid-Piacesian, a little more than three million years ago, and a second mass dispersal as a result of climatic changes that began at the Pliocene–Pleistocene boundary.
The Canis event. The term ‘wolf event’ was proposed by Azzaroli [
16] for the faunal turnover, related to the appearance of the
Canis genus in Europe. This event has been discussed repeatedly in a number of recent studies (see Torre et al. [
17]; Turner [
59]; Spassov [
11]; Sotnikova, Rook [
7]; Rook and Martínez-Navarro, [
4]; Palombo [
27]; Bartolini-Lucenti, Spassov [
60]; Iannucci et al. [
30]; Iannucci, Sardella [
42]; and references therein). The accumulation of data and analysis on this issue shows that there is some diachronism of this phenomenon in Europe, and that it makes sense to distinguish the phenomena of the first appearance of the genus from its mass appearance (see also: Iannucci et al. [
31]).
The earliest appearance of the genus
Canis in Europe: Lacombat et al. [
61] mention several fragmentary remains of mandibles from Vialette, whose age is estimated (see there) to be 3.14 Ma and refer them to
Canis sp. (
Canis has been mentioned in this Late Pliocene locality since Heintz et al. [
62]). Following the publication of Lacombat et al. [
61], the first occurrence of the genus
Canis in Europe is generally accepted to be associated with this Late Pliocene locality (Sotnikova, Rook [
7]; Rook and Martínez-Navarro, [
4]; Palombo [
27]). Iannucci et al. [
31], however, note the existence of chronological heterogeneity of the fauna preserved in the old collections (Crozatier Museum) labeled as Vialette. At the same time, we [
1] (Spassov in Böhme et al.) expressed the opinion that the canid from Vialette most likely represents the genus
Eucyon, known in Europe from the latest Miocene until the Pleistocene beginning. The most informative is the mandibular fragment (2003-5-401-VIA) with m1-m2. The analysis of a cast, kindly provided to me (2006) by A. Monguillon ( Université de Lyon), enabled me to draw some conclusions. The canid is really large, and its dental dimensions are similar to those of a coyote (Lm
1 = 21 mm; p4 = 11.7 × 5 mm; p3 = 10.2 × 4 mm, after cast). This is also reminiscent of the possible
Eucyon—“Canis”
michauxi (Martin [
63]). The lower carnassial tooth of this enigmatic canid from the Pliocene of Perpignan is not known, but the preserved p3-p4 (incorrectly represented in Martin’s [
63] figure as p2-p3) are even larger. The talonid of m1 of the specimen 2003-5-401-VIA is heavily eroded and of incomplete relief, but as seen in
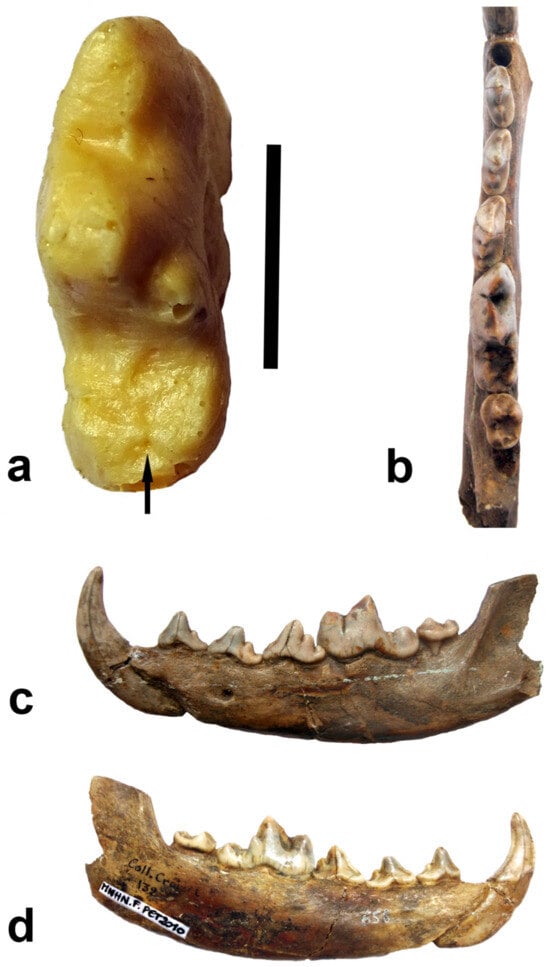
Figure 1, the hypoconid and the entoconid do not contact at their bases. This morphology should correspond to the state of the talonid in Eucyon; in
Canis well-developed cristids that contact and fuse at their bases descend from the hypoconid and the entoconid towards each other (Tedford and Qiu [
64]).
A potential candidate for the earliest known
Canis s. str. (for
Canis (
Xenocyon) arrival in Europe see below)) from Europe is apparently the left semimandible, designated as
Canis neschersensis Croizet et Jobert (see Blainville: Ostéographie, Canis, pl. XIII). It is stored in the Laboratory of Paleontology at the National Museum of Natural History, Paris under number MNHN.F.PET2010 from Perrier-Etouaires (
Figure 1). This number corresponds to an older number from the laboratory of Comparative Anatomy of the Museum—A.C. 658. The museum catalog noted that the find was from Puy-de-Dôme, Auvergne (Puy-de-Dôme = plateau of Perrier = plateau of Boulade, nota mea: NS), specifically from the volcanic sands of Neschers, and was registered under no. 132 in the catalog of the paleontological collection created by l’Abbé Croizet and brought to the museum in Paris in 1839. A number of paleontological remains found in the volcanic sands of the Perrier Plateau have been well-known since the first half of the 19th century. The attribution of the find to the Etouaires site (Ravin des Etouaires = Perrier-Etouaires) in the latest catalogue of the Paris Museum (see above) should be explained by the grouping made by a number of later authors of some smaller sites in the vicinity to the Ravin des Etouaires and uniting them under the name Les Etouaires, assuming that these remains with similar fossilization are also roughly similar in age (Heintz [
65]). Heintz [
65] included in this group of localities the locality of Neschers from the volcanic sands of the region (distinguishing it from the Late Pleistocene Neschers fossiliferous formation), at the same time separating these localities from other and younger Perrier plateau sites such as Roca-Neyra, Pardines, and Peyrolles. Nomade et al. [
19] provided an age of 2.78 Ma for the pumices of la Côte d’Ardé, which they considered stratigraphically very close to Les Etouaires classical site. At the same time, as mentioned above, the fauna from the old collections marked “Les Etouaires” is to one degree or another composite and includes fossils from other localities, as well. That is why I accept the opinion (Iannucci, Sardella [
39]) that the fauna of Les Etouaires, as a whole, is constrained between a plinian fall dated at ca. 3.11 Ma and an overlying debris avalanche, whose pumices (contemporary to the Roca-Neyra fauna) have been dated (Nomade et al. [
19]) at ca. 2.60 Ma. As a result, I can agree with the statement that the
C. neschersensis find should fall, in general, within this time interval (Iannucci et al. [
31]). After A. Iannucci (pers. comm.), the mandible of
C. neschersensis has a different fossilization than some findings of certain Pliocene age (like the suids) from Perrier. It could be coeval with the remains of
C. (
Xenocyon)
falconeri from Boulade (also from the Perrier plateau), a locality close to Roca-Neyra and correlated with it in age (2.6 Ma) (Bartolini-Lucenti, Spassov [
60]), or it could be younger (?), but it seems to me that it is most likely earlier (see above; Heintz [
65]) and could refer to the end of the early Villafranchian (MNQ16).
Following is a short comparative description of MNHN.F.PET2010: measurements: Lp1-m3 (alveolar) = 76.5 mm; m1 = 23.5 × 8.5; mandible height under m1 (labially) = 21.2. The size is slightly inferior to
C. etruscus—
C.
arnensis and closer to “
C. apolloniensis.” The mandible is distinct from the Villafranchian group of taxa after several morphological characters, namely by the specific premolar features: large premolars, strongly developed on all pm (p2-p4) posterior additional cuspids (strong posterior additional cuspid on p2 including a very strong secondary additional cuspid on p4), and a lack of any gap between them (p4 partially overlaps the base of the anterior paraconid ridge). It is also distinct by some of its plesiomorphic (not wolf-like) characteristics, such as the narrow m1 trigonid with a rather flattened labial surface, the well-developed pre-hypoconid and pre-entoconid on the m1 talonid, and the strongly convex ventral surface of the mandible corpus. The typical
Canis cristids on the talonid cuspids are well-developed, but the entoconid is narrow and mesiodistally elongated. The m1 talonid is relatively large. The M
2 is relatively short, but broad. Its mesial cuspids show a plesiomorphic condition: They are almost equally large, positioned in a straight line almost perpendicular to the longitudinal axis of the teeth (i.e., the labial one is not mesially placed in relation to the lingual one) (Spassov [
66]). The teeth show, on the other hand, some derived characteristics that resemble
C. mosbachensis, which appeared in Europe towards the end of the early Pleistocene: the paraconid of m1 is oblique and exceeds the height of p4, and p3 is relatively low-set in relation to adjacent premolars. Therefore, a more detailed comparison with this species makes sense: The mandible from Neschers differs from the European representatives of
C. mosbachensis (as well as from all Villafranchian European
Canis, as I already noted) by the remarkably short alveolar space, which is why there are no gaps between c1-m2 (due to lack of space, m2 is very high positioned on the surface of the mandible ramus); the labial outline of m1 is flat; the line of the p4 base is horizontal, in an angle with the line of the bases of p2-p3; p2 is with a well-marked posterior additional cuspid, which appears to be absent in the European representatives of the species and very rare in the Asian subspecies; and the mandible corpus is (as noted above) with a strongly convex ventral surface. It is interesting that in some features, such as the oblique m1 paraconid, the height of paraconid of m1 exceeding the height of p4, the relatively convex mandible body, the lack of gaps between teeth, the shape of m2, the position of p4 base in relation of the bases of p2-p3, the strong posterior additional cuspid in p2, the strong secondary additional cuspid in p4, the complex m1 talonid relief, and the mesio-distaly elongated m1 entoconid, the mandible of the
Canis from Neschers resembles
Canis chihliensis palmidens (NIH 164, Paris Museum) from Nihewan, China, from the Pleistocene beginning (2.4 Ma). In summary, it can be said that the unclear geological age and the lack of more material, bearing in mind the not small variability in
Canis, do not give certainty when discussing
C. neschersensis as the earliest
Canis in Europe. However, some arguments noted above regarding its probable age (possibly between 2.8 and 2.6/2.4 Ma?), as well as its specific morphology, give reason to also mention, in the discussion of the early
Canis appearance, this fossil as the first putative representative of the genus
Canis in Europe.
Torre [
67] and Iannucci et al. [
31] noted that the presence of
Canis (as probable “
Canis aff.
etruscus”) from “Etouaires” was reported also by Heintz et al. [
62] based on fossil material from the Bravard Collection housed in the British Museum and listed by Lydekker with provenance as “Tour de Boulade.” Torre [
67] (following Lydekker) considers that this fossil must “belong to a large wolf which is very similar to those of the last Glaciation.” Here we have to note that from Boulade (near Roca-Neyra), the earliest European remains of
Canis (
Xenocyon) are described based on old collections that are now kept at the Claude Bernard University in Lyon (Bartolini-Lucenti, Spassov [
60]). The tower “Tour de Boulade” itself is in the same region, above the Parentignat bridge, but at about 4.5 km from the mentioned site of Boulade. It is not clear whether the old designation of the site as “Boulade” has anything to do with “Tour de Boulade” (where Late Pleistocene fauna was indicated; Fosse et al. [
68]). At the same time, however, as Sebastien Nomade clarifies (in a letter to S. Bartolini-Lucenti, 2021), the site La Boulade, where the remains of the mentioned
C. (
Xenocyon)
falconeri should come from, is about 2.5 km away from Roca-Neyra, and stratigraphically and geochronologically, La Boulade and Roca-Neyra are both dated between 2.59 and 2.60 Ma. A comparison of a photograph from the cast of the mandible, stored in the collections of the British Museum (owned by the University of Florence and kindly provided to me by Saverio Bartolini-Lucenti) shows the following: The mandible is larger overall than a recent European
C. lupus and has more elongated m2, but at the same time, shows considerable similarities with the wolf. From
C. (Xenocyon)
falconeri (IGF 865, coll. of the University of Florence), it is distinguished by the more robust m1 (especially the paraconid part); the shorter premolar row and smaller p1; and the larger i3. The articular process is wolf-like in shape, not as in
C. (
Xenocyon), and is rather more robust. Evolved wolves reach (albeit much later in time) the apomorphic features of the carnassial tooth of
C. (
X.)
falconeri. However, the m1 hypoconid of the mentioned mandible is not as centrally positioned on the talonid and is not as significantly larger than the entoconid, as it is in a
C. (
X.)
falconeri sample from Boulade (Roca-Neyra).
In relation to the time of the first appearance of
Canis in Europe, some other unclear cases deserve to be noted. Among them are a mandible from Csarnota final Pliocene (Hungary), and a mandible fragment with p3 and p4 is mentioned as “
Canis sp.” in Capeni (Radulescu, Samson [
55]; see also above for the remains of
Equus also noted in this locality). At the same time, Marciszak et al. [
69] mentioned, but did not describe the presence of
Canis in the Late Pliocene Weze 2 locality (ca. 2.8–2.6 Ma). Argant [
70] reported from Saint Vallier (with a probable age of almost 2.5 Ma; Nomade et al. [
19]) the presence of scarce long bone fragments and an occipital skull fragment that resembled
Canis. At the same time, the presence of “
Canis” (?) is also established in Chilhac (Monguillon-Douillet A., [
71]), whose age is determined at ca. 2.36 Ma (Nomade et al. [
19]). In relation to all this, we must not forget the presence of
Canis at Neschers. But it seems more likely to me that the remains (or most of them) refer to
Eucyon, which must have been displaced by
Canis in its later mass appearance in Coste San Giaccomo, Slivnitsa, and Senèze, than to belong to
Canis s. str. Three separate teeth from the close-in-age locality of Varshets, Bulgaria (MNQ17, ca. 2.5 Ma), have been assigned to this genus (Spassov [
11]). These cases cannot be resolved without a special, additional comparison.
- 2.
The mass appearance of the genus: In Western Europe (France, Italy, Spain), the
Canis mass appearance occurred in the period of about 2.2–1.98 Ma (Palombo [
27]), but this can be said for all of Europe in general. We can call this dispersal, using Azzaroli’s concept [
16],
Canis event s. str. or mass
Canis appearance in Europe. The earliest
Canis mass arrival on the continent must be related to the Coste San Giacomo unit. We can claim that the following localities (starting from east to west) record practically the same wave of dispersal of the “wolf-like”
Canis to Europe: Slivnitsa, Bulgaria (see below: Spassov [
11,
72]), Coste San Giacomo (2.2 Ma: Florindo et al. [
73]) and Quercia (2.2–2.1 Ma: Iannucci [
74], Italy), and Senèze, France (its base age is of ca. 2.2 and the youngest fossils ranged in age between 2.10 and 2.08 Ma after a new investigation in. prep.: Eric Delson, pers. comm.). It seems that these localities, which record the first secure mass invasion of the genus (Spassov [
11,
72,
75]; Rook, Martἱnez-Navarro [
4]; Iannucci [
74]) have a very similar/close age and should be placed in the C. San Giacomo Unit. As it seems, two species entered practically simultaneously from the East to Europe (
C. etruscus and
C. arnensis are apparently present simultaneously in the locality of Slivnitsa; Spassov [
72]), and two species of
Canis are detected in Senèze after a new investigation (chapter in press on Senèze carnivores: A. Argant, pers. comm.). Slivnitsa and Senèze have very similar fauna and should be of the same/very close age (Spassov [
75]). They show other mass dispersals, as well (see below). The Slivnitsa faunal event must, therefore, correlate (Spassov [
34]) with the climatochronologic zone SCT10 of Zubakov and Borzenkova [
76]. This zone, documented in Georgia and the Azov region, has an estimated age predating the Olduvai warming (between the Reunion subchron [2.193 Ma] and the beginning of the Olduvai event [ca. 1.95 Ma]).
Canis (assigned to
C. cf.
senesensis) is also mentioned for the Khapry faunal complex, southern Russia (especially for the Liventsovka and Khapry s. str. localities; see Titov [
77]). After the statement of several authors (e.g., Sotnikova et al. [
78]; Sotnikova, Rook [
7]) these
Canis finds must be correlated biostratigraphically to MNQ17, and along the Northern Black Sea coast, the genus should have already appeared in the Middle Villafranchian (MNQ17). Spassov [
11] cast doubt on the dating of these finds, suggesting that they could originate from the upper and younger faunal levels of the multilayer Liventsovka section. The occurrence of three and possibly six equid
Equus species also suggests a heterogeneity in age of the Khaprovskiy (Khapry) complex (Eisenmann [
53]). Titov [
77] estimated that the Khapry Sands, which extend for more than 120 km and up to 2 km width, could have accumulated for as long as 400,000 years and have an age from the early MNQ17 (excluding the earliest Middle Villafranchian) to the pre-Olduvai part of the Matuyama chron, between 2.6/2.5 and 2.2 Ma. According to Iannucci and Sardella [
42], the occurrence of
Pachycrocuta brevirostris there makes even wider the time limits during which the sands of the Khapry complex (having negative magnetic polarity and, thus, a pre-Olduvai age) were accumulated (between 2.6 and not less than 2.0 Ma). Considering the possible upper limit of the age of the Khapry complex (2.2–2.1/2.0 Ma), it is very likely that the appearance of
Canis there also coincides with the C. San Giacomo Faunal Unit and represents a manifestation of the same
Canis event traced west of the Black Sea in that time (Spassov [
11]; see also below). Ultimately, even if the mass
Canis arrival took place in the Azov region in the late NMQ17 (?), to the west of the Azov Sea, this penetration was recorded at the geologic time of Slivnitsa and Senèze (the Coste San Giacomo Unit).
Large felid events (Acinonyx and Puma dispersal in the latest Pliocene [MNQ16] and the Panthera dispersal in the Early Pleistocene [beginning of the Late Villafranchian—MNQ18a]).Acinonyx is thought to have originated in Africa, where the oldest remains (3.85–3.60 Ma) are likely from Laetoli (Werdelin, Dehghani [
79]). The oldest remains of Eurasian
Acinonyx are known from the MN16 of Perrier (Puy de Dôme), where the genus and probably the species A.
pardinensis was found in two different, but very close sites: La Côte d’Ardé (with
A. p. pardinensis) and Les Étouaires (with
A. p. arvernensis), the former being the type locality according to Hemmer et al. [
80]. Interestingly, the new dating of La Côte d’Ardé, by which Nomade et al. [
19] dated, in fact, the very nearby site of Les Étouaires s. str., is 2.78 Ma. This means that the two disputed taxa are practically of the same age (above, we drew attention to the assemblage of fauna from different localities, often attributed to the Les Étouaires locality). This casts doubt on the subspecific status of these taxa. It seems that the fossil Eurasian cheetah is known from a large number of localities from the Early Villafranchian (only in Europe) until the Middle Pleistocene and from S.-W. Europe to China (Cherin et al. [
81]).
The Les Étouaires locality (s. str.?) also seems to be associated with the earliest European record of the Eurasian
Puma pardoides and probably also with the earliest saber-toothed cat
Megantereon (Hugueney et al. [
82]; Hemmer et al. [
83]; Hemmer, Kahlke [
84]; Cherin et al. [
85]). The Eurasian fossil
Puma is found with certainty in more than 10 Villafranchian localities known from S.-W. Europe till Mongolia (Cherin et al. [
85]). Hemmer et al. [
83] suggested, based on some fossil material, that the genus originated in Africa (Laetoli), but this suggestion was not supported by the analyses of Werdelin, Dehghani [
79]. The origin of the genus is more likely to be related to Central Asia, and its earliest known occurrence (3.1 Ma) appears to be in Shamar, Northern Mongolia (Sotnikova [
86]; Hemmer [
80]). According to the available data, both
Puma and
Acinonyx appeared in Europe at the very end of the Pliocene (MNQ16b) in the Perrier localities (“Les Etouaires”).
Much later, the genus
Panthera appeared in Eastern Europe. The oldest finds of the genus are probably those from the Laetoli upper unit (3.85–3.60 Ma) (
Panthera sp.; see Werdelin, Dehghani [
79]). After Hemmer and Kahlke [
84], the earliest Eurasian “jaguars” (sensu Hemmer [
87]) (i.e., the early representatives of
Panthera in Eurasia) obviously descended from an African
Panthera population. The same author believes that the Eurasian fossil representatives of the genus do not differ significantly from the modern jaguar and should be included in the same species [
84,
88] (Hemmer [
88]; Hemmer, Kahlke [
84]; and references therein). This opinion is not accepted by all authors (see Jiangzuo [
89]), and until recently, the question about the taxonomic appartenance of the Villafranchian Eurasian
Panthera representatives remained controversial. For a clearer characterization of phylogenetic lines, we can enrich the taxon
Panthera with one more clade, the subgenus
Onca, in which jaguar-like cats can be divided into
P. (
Onca)
toscana—the earliest (Late Villafranchian) Eurasian fossil jaguars;
P. (
Onca)
gombaszogensis—the later representatives of Eurasian jaguars; and
P. (
Onca)
onca—the American jaguars (for the correct transcription of the name as
P. gombaszogensis, not
P. gombaszoegensis, according to ICZN rules, see Wagner [
90]).
The smallest and most ancient form of the European jaguar
P. (
Onca)
gombaszogensis toscana is recorded, apparently, for the first time in Slivnitsa, Bulgaria (Spassov [
11,
72]).
P. gombaszogensis has also been recorded in the Villafranchian fauna of Tegelen, but the Tegelen large mammal fauna probably does not represent a single time frame, and a large part of the fauna originates, it seems, from sediments deposited a little later, during the Olduvai Subchrone (Hoek Ostende [
91]). The presence of the species (see Iannucci et al. [
31]) at Puebla de Valverde (2.13–1.98 Ma) is uncertain. A little later than the appearance in Slivnitsa is the finding of
P. gombaszogensis in localities such as Gerakarou 1, Greece (Koufos [
92]), and in Olivola, Italy (Torre et al. [
17]; Gliozzi et al. [
18]; Iannucci et al. [
31]). Gerakarou 1, where
Pachycrocuta brevirostris is also present, should have an age later than Slivnitsa and related to the boundary between the C. San. Giacomo Unit (MNQ18a) and the Olivola Unit (MNQ18 b) (Spassov [
11]; see
Section 2.2. and
Section 2.3).
Pachycrocuta brevirostris appearance. The first appearance of this giant hyena in Europe is considered by a number of authors as a faunal event because of the high impact of this giant supercarrion eater hyaenid in the Early Pleistocene faunal assemblages, but often also because of the “simultaneous” appearance with another hipercarnivore,
Panthera gombaszogensis, in Europe (Martínez-Navarro [
93]; Rook, L., Martínez-Navarro [
4]; Croitor et al. [
94]; Iannucci et al. [
31]). The dispersal of this felid, however, is earlier, as noted above. At the same time, other species noted their mass distribution in Eastern or Western Europe at that time (see
Section 2.2). The so-called
Pachycrocuta brevirostris event is frequently considered to mark the beginning of the Late Villafranchian [
4,
93,
95] (Martínez-Navarro [
93]; Rook and Martínez-Navarro [
4]; Sianis et al. [
95]), but according to the understanding expressed in this study, this beginning is marked by the C. San Giacomo-Slivnitsa–Senèze turnover (see
Section 2.2). One of the earliest occurrences of
Pachycrocuta in Europe is likely from the Iberian Peninsula: Based on some
P. brevirostris remains from the localities P-1 and SCC-1 (~2.12–1.92 Ma) in the Fonelas area, as well as from the locality of Almenara-Casablanca 1 (pre-Olduvai age), Madurell-Malapeira et al. [
96] have placed the
P. brevirostris event before the Olivola Faunal Unit. These rare finds, however, apart from probably being close in age to the beginning of the Olivola FU, are poorly informative and can generally be considered rather tentatively (Iannucci et al. [
30]). The giant hyena has been recorded in Gerakarou-1, Greece (Koufos [
92,
97]), and its presence in this locality must be among the first appearances of this carnivore on the continent (Spassov [
11]; Iannucci et al. [
30]). The age of Gerakarou should be approximately at the boundary between the C. San Giacomo and Olivola faunal units, slightly later than that of Slivnitsa (which we include in Coste San Giacomo FU) and similar to the very beginning of Olivola FU (Spassov [
11]). In Olivola, Italy,
P. brevirostris is well-known (Gliozzi et al. [
18]; Iannucci et al. [
30]) and pointed out as one of the main faunal elements initiating the Late Villafranchian faunal dispersal events (Azzaroli [
9]). The Olivola locality has not been directly dated. The lowest possible placement of Olivola is around 2.1 Ma (Napoleone et al. [
98]), and it is usually placed between 2.1–1.9 Ma (~2 Ma: Iannucci et al. [
31]). Ultimately, the beginning of the
Pachycrocuta brevirostris event in Europe should be dated, according to existing data (at least west of the Azov Sea), to about 2 or a slightly more than 2 Ma (it is not clear whether the species appeared in Khapry, in the Azov region, shortly before).
Hippopotamus event. The dispersal of hippos into Europe is considered as an event of biostratigraphic importance and related to climatic changes (Rook, L., Martínez-Navarro [
4]). The first appearanceof the
Hippopotamus in Europe is now attested at 2.2 Ma (Coste San Giaccomo, Italy; see Bellucci et al. [
99]; Fidalgo et al. [
100]; Iannucci et al. [
31]) and is possibly also from Elis (Greece), with unclear age. The age of Elis is mentioned as “Middle Villafranchian” (?) (Reimann, Strauch [
101]) and “Lowest Pleistocene,” at ~2.1 Ma (Athanassiou [
102]; Athanassios Athanassiou, pers. comm.). The
Hippopotamus incisor from Coste San Giacomo was collected during field collection, ex situ, causing some authors to doubt the age of the finding (Marra et al. [
103]; Mecozzi [
104]). However, field activities and excavations at Coste San Giacomo have pointed out that the vertebrate assemblage comes from a single fossiliferous level (Iannucci et al. [
31]). After Mazza and Rustioni [
105], a first phalange from Senèze (FSL 211082), previously ascribed to
Equus sp., must be attributed to
Hippopotamus, a claim based on which Iannucci et al. [
31] accept the presence of the hippopotamus (also mentioned in the older literature) also at Senèze. However, two posterior first phalanges cataloged as FSL 211,082 are discussed as
Allohippus senezensis by Eisenmann and Delson in a publication currently in preparation (E. Delson, pers. comm.) In sum, it seems that there is no evidence of the
Hippopotamus in the Senèze assemblage.
The question of how, from where, and under what climatic conditions the
Hippopotamus colonized Europe is of interest. Despite their aquatic lifestyle, hippopotamuses cannot swim (Mazza [
106]). There are no reliable data for a land bridge via the Gibraltar Straits or via Sicilia during the Villafranchian (Spassov [
34] and references therein), which means that the hippo most likely could not have passed from North Africa to Europe via these hypothetical dispersal routes (especially Gibraltar) discussed by a number of authors (see Fidalgo et al. [
100] and references therein). The logical route of dispersal, also supported by the earliest finds in the area, is that via the Levantine corridor (see Made et al. [
107]; Fidalgo et al. [
100]). The route from the east to the continent is also the route of almost all migrations of large mammals to Europe (Spassov [
11,
34]). This entry route for the hippopotamus is supported by the registered Meria cooling documented, as already mentioned, in the Black Sea region in the time between the Reunion subchron and the beginning of the Olduvai event showing signs of a temporary closing of the Bosphorus (Zubakov and Borzenkova [
76]).
Iannucci et al. [
31] note that the appearance of the hippopotamus in Europe documented an African dispersal of a species linked to humid conditions in a context that is generally deemed to denote the spread of open-adapted faunal elements of mainly Asian affinities. But did the appearance of the hippopotamus really occur in humid conditions at that time on the continent, and does this appearance really contradict the spread of open-adapted faunal elements from the East to Europe? In fact, the hippo cannot stand the strong sun and spends most of the day in water to stay cool and hydrated. At the same time, the presence of water bodies does not mean wet conditions. The tropical savannah conditions in which
H. amphibius lives today are characterized as usually being very dry. This animal does not need humid conditions, but rivers and patches of water scattered throughout in open landscapes (Parker [
108]), conditions that do not contradict a relatively cool environment in southern Europe ~ 2.2 Ma ago.
The Bison (Eobison)—B. (Bison) replacement and the Lower Epivillafranchian boundary. The end of the Villafranchian shows, according to a number of researchers, a transitional fauna between the Villafranchian one and that which is characteristic of the Middle Pleistocene. This episode in the history of the megafauna is referred to in the literature in different ways: Latest Villafranchian sensu Koufos, [
109], Final Villafranchian sensu Spassov [
11], and the term Epivillafranchian, proposed at the beginning of the 1960s, which has acquired the widest distribution and gradually become established as the name of this biochron (Kahlke [
110]; Kahlke et al. [
37] and references therein). Its time span is between the Late Villafranchian s. str. and the Galerian biochrons (1.2–0.9 Ma). In general, it is defined as the time between the
Praemegaceros verticornis—Bison menneri first occurrence and the
Crocuta crocuta first occurrence] (Kahlke [
110]; Bellucci et al. [
111] and references therein). The disappearance of the primitive bison
Bison (
Eobison) and its replacement by the evolved bison of the subgenus
B. (
Bison) seems to be among the important faunal events reflecting climatic and landscape changes (van Asperen, Kahlke [
2]) and, thus, indicating the boundary between the Late Villafranchian and the Epivillafranchian.
The putative ancestor of the bison is
Leptobos (Cherin et al. [
112] and references therein), or it is its possible sister species, bearing in mind the new data (Akbar Khan et al. [
113]) about the rather early appearance of the primitive bison. While
Leptobos has been present in Europe since at least 3.3 Ma and throughout the Villafranchian mammal age (Masini et al. [
114]; Mead et al. [
115]),
Bison probably appeared in Asia and entered Europe much later. Its first occurrence is registered in the Upper Siwaliks of Pakistan with
Bison (
Eobison) cf.
sivalensis in the Early Villafranchian, between 3.3–2.6 Ma (Akbar Khan et al. [
113]) and reaching the boundaries of Europe (Dmanisi) ~1.76–1.8 Ma ago with
B. (
E.)
georgicus (Burchak-Abramovich and Vekua, [
116]; Sorbelli et al. [
117]). The last primitive bison that should be included in the subgenus
B. (
Eobison) are known from Pirro, Italy (
B. degiulii), and Apollonia, Greece, (
B. cf.
degiulii) (with probable ages of about 1.3 and about 1.2 Ma). They are distinguished by an increasing size compared to earlier representatives of the subgenus and by more massive metapodials (Sorbelli et al. [
117]). The same authors note that the progressive climatic deterioration and the spread of open environments led to an increase in metapodial stoutness and in the body size of these forms. Increasing size is a natural process in the evolution of many taxa, but it is difficult to agree that the massiveness of the metapodials of these bisons is due to inhabitance in more open and arid landscapes. It has long been known, as a result of a number of studies, that active running in steppe landscapes and on terrains harder than forest landscapes lead to the exact opposite result—to a reduction in the massiveness and elongation of the metapodials, both in equids and in artiodactyls (Gromova [
118]; Eisenmann [
119]; Scott [
120]). The massiveness of the metapodials of the last representatives of
B. (
Eobison) should be due to the increase in size and, hence, the weight of these forms, which at the intensification of the cursoriality in open, plain landscapes, leads for biomechanical reasons to the shortening of metapodials (Gambaryan [
121]), and as a consequence of this, to a change in their proportions and an increase in their massiveness.
2.2. Major Villafranchian Events. A Summary
The data on the first appearance of different taxa in the Villafranchian of Europe are constantly replenished, and today, it is clear that the processes are more complex and often diachronic (Iannucci et al. [
31]). There is hardly any doubt that the main driving forces of mass dispersals are more significant climatic and, hence, environmental changes. Some of the more significant dispersals are briefly summarized below.
End Pliocene (Early Villafranchian: MNQ16) events. As noted above, the time from the Mid-Piacenzian (~3.3–3.0 Ma) marks the beginning of a particularly intense dispersal of species, of a new faunal exchange between Eurasia and Africa, as well as of the entry of new mammals into Europe from the East in the Early Villafranchian (for the first appearances of individual faunal elements, see above). At that time, Leptobos appeared in Southern Europe, and this seems to be the time of origin of the genus. The appearance of Mammuthus in Europe is associated with this time. Its earliest finds are usually associated with the species M. rumanus, and its first secure occurrence (Romania and Moldova) is about 3.2 Ma ago. The first appearance of the monodactyl horses in Europe is a question we cannot yet accept as definitively resolved. It is possible that there was a first (unsuccessful?) dispersal on the continent in the Mid-Piacenzian, a little more than three million years ago, and a subsequent mass dispersal during climatic changes that began at the Pliocene–Pleistocene boundary.
The site of Les Étouaires marks the first occurrence of several taxa of large carnivores. The “Les Étouaires” faunal assemblage is, as it seems, complex and made of various outcrops and levels. They do, however, appear to be relatively close in age within the MNQ16 zone (see above). The earliest remains of
Acinonyx pardinensis are from Les Étouaires and from the neighboring La Côte d’Ardé site, accurately dated after the deposits from the base of the Les Étouaires fluvial sequence at Côte d’Ardé (2.78 Ma after Nomade et al. [
18]). The Les Étouaires locality itself is also correlated with the age of these deposits (see the corresponding passage above). From these levels also appear to be the remains of the earliest
Puma in Europe. The first occurrence of
Canis s. str. (possibly with
C. neschersensis) is not entirely clear. The age of the type locality of “Neschers” is not clear, but we have some reason, as we noted earlier, to assume an age close to Étouaires. This locality also appears to be associated with the first (certain) occurrence of
Pliotragus ardeus (Cregut-Bonnoure [
22]; Croitor et al. [
94]). Its occurrence in localities of earlier age, for example, in the Late Miocene of Stratzing, Austria, can most likely be explained by the presence of higher stratigraphical levels there (Gentry [
122]).
The Quaternary beginning and the Middle Villafranchian faunal events. The first certain appearance of the monodactyl horses (
Equus or
Allohypus, according to different taxonomic interpretations) is related to the Pliocene/Pleistocene boundary. A number of localities mentioned above, and with an age very close to the beginning of the Quaternary (close to or synchronous with the Gauss–Matuyama transition (i.e., 2.588 Ma), show the first certain mass appearance of the horse. As such, these mass dispersal event localities, such as Roca-Neyra, for example (2.60 Ma) should mark the beginning of the Middle Villafranchian and the beginning of MNQ17, not the end of MNQ16 and the Early Villafranchian. A little later, around the beginning of the Quaternary, the suid
Sus strozzi appeared in Europe, commonly found there in the first part of the Early Pleistocene of Europe (Cherin et al. [
123]). Of all the earliest finds, the most securely dated seems to be that of Saint-Vallier (Iannucci et al. [
31]), a type locality of the MNQ17 zone, with an age older than 2.4 Ma, perhaps close to 2.5 Ma (Nomade et al. [
19]). It seems that the Pliocene/Pleistocene boundary is also associated with the first occurrence of
C. (
Xenocyon)
falconeri in Europe, according to remains from France with a probable age of 2.6 Ma (Bartolini-Lucenti, Spassov [
60]), and from Poland at 2.4–2.2 Ma (Marciszak [
69]). The replacement of the Archaic
Mammuthus rumanus with
M. meridionalis probably also occurred soon after the beginning of the Quaternary and in the Middle Villafranchian: The earliest
M. meridionalis apparently appear in Khapry, S. Russia (Titov [
77]).
Slivnitsa–Coste San Giacomo–Senèze turnover. Although the appearance of
Canis in Europe takes place probably, as we noted already, at the end of the Pliocene (or around the Pliocene-Pleistocene boundary), the mass appearance of the “wolf-like”
Canis s. str., which we can associate with the “wolf event” of Azzaroli [
16], becomes later. This wave of dispersal from the East is associated with the time between 2.2–2.1 Ma. Coste San Giacomo (CSG), Italy (Napoleone et al. [
98]), is associated with an age of 2.2 Ma. The base age of Senèze (France) was also found to be ca. 2.20 Ma after a new investigation (in prep.: E. Delson, pers. comm.) (2.21 to 2.09 after Nomade et al. [
19]). In CSG, the gomphothere
Anancus (relic from there?) is still found, but in both localities, as well as in the apparently similar in age Slivnitsa (Bulgaria), “wolf-like
Canis” appear, such as
C. arnensis, which is similar in size to the coyote (Brugal, Boudadi-Maligne [
124]) or slightly larger. Two species of Canis are present in Senèze, according to the findings from a new investigation (chapter in prep. on Senèze carnivores: A. Argant, pers. comm.), and apparently also in Slivnitsa (Spassov [
11,
72]). We correlate the Slivnitsa faunal event (Spassov [
34]) with the climatochronologic zone SCT10 of Zubakov and Borzenkova [
76]. This zone, documented in Georgia and the Azov region, has an estimated age between the Reunion subchron and the beginning of the Olduvai event. During this period, which corresponds to the Meria cooling documented in the Black Sea region, there are indications that some Aegean islands close to the Anatolian coast were connected with Asia Minor (Dermitzakis [
125]) and that the Black Sea was a freshwater sea; this suggests a temporary closing of the Bosphorus (Zubakov and Borzenkova [
76]). This explains the new wave of faunal dispersal from the East during that time. With Slivnitsa, a Villafranchian Balkan faunal unit can be designated, but it would be most logical that Senèze and Slivnitsa, together with Coste San Giacomo, should be united in a common faunal unit (CSG Unit) and separated in the MNQ18a zone (sensu Spassov [
11,
34,
126]), thus marking the beginning of the Late Villafranchian. The MNQ18a zone and CSG unit in the range indicated above are characterized in part by the mass appearance of
Canis and by the first appearance of
Panthera on the continent (Slivnitsa), by the first appearance of
Ovis and
Hemitragus (Slivnitsa and Senèze) (Spassov [
11,
34,
72,
75]), as well as by the first appearance of the hippo in Coste San Giacomo (Bellucci et al. [
99]; Fidalgo et al. [
100]). The dominance of bovid species over cervids (Slivnitsa) at that time is also an indication of the cooling and opening of the landscape.
In the context of the above, some data and ideas about the earliest possible entry of the human genus into Europe are of interest. In recent years, an increasing number of discoveries have supported the idea that the earliest human occupation of Europe was via the Bosphorus/Peri-Pontic pathway only (Strait et al. [
127]) and took place during the Villafranchian, significantly predating 1 Ma. The time of the earliest human appearance in Europe could be related to conditions of increasing aridification and to a domination of open/mosaic landscapes.
The time of Slivnitsa (Meria cooling), which should be the time interval between the Reunion subchron and the beginning of the Olduvaian event, has been proposed for the time of the earliest possible appearance of the genus
Homo on the continent (Spassov [
34] and references therein). The discovery of lithic artefacts at the site of Kermek (Azov Sea region, S. Russia) with an age of ca. 2 Ma (Shchelinsky et al. [
128]; M. Gurova, Inst. of Archaeology, Sofia, pers. comm.) seems to confirm the above hypothesis about the time of the first (probably unsuccessful) appearance of
Homo in Europe.
The Olivola FU beginning. The age of the Italian locality of Olivola, which provided the name of the Olivola Faunal Unit (Gliozzi et al. [
18]), has not been directly dated and is usually placed at ca. 2 Ma; its lowest possible placement is around 2.1 Ma (Napoleone et al. [
98]). The very beginning of the Olivola Faunal Unit (the Coste San Giacomo/Olivola FU boundary) is related to the appearance of
Pachycrocuta in Europe and the wide spread of
Canis. The first occurrence of
Pachycrocuta seems to be immediately after that of the mass appearance of
Canis in the Coste San Giacomo FU. Apparently, the appearance and spread of these carnivores is related to the dispersal of a number of ungulates, as a result of the change (the opening) of the landscape. After some investigations (Croitor et al. [
94]), the dominant ruminant species in Eastern Europe (after the example of the Dacian Basin) before the
Pachycrocuta event (
Pliotragus ardeus,
Gazellospira torticornis,
Rucervus radulescui,
Metacervocerus rhenanus, and
Mitilanotherium inexspectatum) became extinct during the Pachycrocuta faunal turnover and were replaced by a more cold-adapted assemblage of ruminants (
Megalovis latifrons,
Eucladoceros sp.,
Dama sp., and
Praemegaceros obscurus). The mentioned turnover is also manifested in Western Europe, where the time span of the Olivola FU shows the peak of bovid diversity and the disappearance of
Gazella borbonica (Masini et al. [
114]).
The Late Villafranchian/Epivillafranchian boundary. If we assume that the Epivillafranchian represents a separate episode of the development of the European megafauna, then the end of the Late Villafranchian in a narrow sense should end at the boundary with the Epivillafranchian. The boundary between these two episodes of faunal evolution is marked by new climatic changes leading to a new faunal turnover. Among the many faunal changes (some of which are controversial for taxonomic reasons; Bellucci et al. [
111]), we can mention again the disappearance of
Bison (
Eobison), whose last occurrence is at Pirro and probably also at Apollonia (Sorbelli et al. [
117]), as well as the first occurrence of
Praemegaceros verticornis and
Bison menneri, and perhaps also of
Megaloceros savini (Kahlke [
110]; Bellucci et al. [
111] and references therein). Also worth mentioning here is the appearance of the evolved form of
Sus strozzi (Iannucci [
74]), which is possibly a separate subspecies (considered by some to be an ancient form of the later
Sus scrofa).
2.3. Correlation of the Villafranchian Biochronology between Eastern and Western Europe (Correlation of Selected Eastern European Localities)
The study of the Villafranchian faunistic complexes from Eastern Europe and Central Asia is of great significance for the elucidation of the origin and evolution of this fauna in Europe as a whole. There is a logical trend to correlate stratigraphically the biocomplexes in Eastern and Southeastern Europe and even in Central Asia with those earlier defined in W. Europe by using the same biostratigraphic criteria. The dispersal of Villafranchian faunal elements through Europe is usually an instantaneous event from a geological point of view, but in some cases, W. Europe is reached later, and some species known in Eastern Europe never even reach Western European territories (Vangengeim [
129]; Spassov [
11]; Cregut-Bonnoure [
22]). The environmental differences and the zoogeographic features of the fauna spread from the Urals to the most western and southwestern territories of the continent create a number of difficulties in the biochronological correlation. I have specifically focused on this issue in previous studies, in which I attempted such a correlation (Spassov [
11,
75]). Many new data (the discovery of new paleontological localities, new mass fossil material, and the absolute dating of a number of key deposits) make it possible to refine this correlation (
Figure 2).
Early Villafranchian. The Balkan localities of mammalian megafauna from the early Villafranchian are few. The Romanian localities with
Mammuthus rumanus, Cernãteşti and Tulucesti, as well as the Moldovian locality of Ripa Skortselskaya, must be placed by geochronology at 3.2–3.1 Ma (Radulescu et al. [
55]; Nikiforova et al. [
45]; Andreescu et al. [
38]). The
M. rumanus mandible from Bossilkovtsi (Bulgaria), originating from Pliocene sands (M. Böhme, University of Tubingen: pers. comm.) in its morphological features, is similar to the stage of this species from the indicated Romanian localities, and the Bossilkovtsi locality should be close in age. The archaic
Mammuthus maxilla from Tsotylio, Greece, is also probably of a similar age (Kostopoulos, Koulidou [
49]).
Middle Villafranchian. The Khapry faunal complex (Southern Russia), in which fauna is revealed in at least seven palaeontological localities, has a wide possible temporal span of ~400,000 years and occupies a time interval from the beginning of the Middle Villafranchian (not including its beginning) to ~2.2 Ma (Titov [
77]). Given the presence of
P. brevirostris, we can place this faunal assemblage at approximately between 2.5 and 2.1 Ma (see the discussion in the chapter The
Canis Event). The earliest Middle Villafranchian locality on the Balkans appears to be Varshets, Bulgaria. According to its fauna, the presence of
Nyctereutes tingi (Spassov [11; Tamvakis et al. [
130]), the presence of the primitive
Gazellospira sp. (known in Roca-Neyra) (Spassov [
54]), and the evolutionary stage of
Martes (Marciszak et al. [
131]) shows a transition from the fauna of Roca-Neyra to that of Saint-Vallier (the benchmark locality of the MNQ 17 zone), with which there is a great similarity (Spassov [
11,
34]). Given the age of Roca-Neyra (2.6 Ma) and the assumed age of Saint-Vallier (over 2.4 and under/close to 2.5 Ma) (Nomade et al. [
19]), we can assume that the age of Varshets is around 2.5 Ma. The Dafnero site, with several localities, is a typical MNQ17 site, which is somewhat later than Varshets after its fauna (Koufos, Kostopoulos [
29,
132]; Spassov [
11]), which is confirmed by the results obtained for its absolute age (2.4–2.3 Ma, probably closer to 2.3) (Benammi et al. [
133]). The Greek localities Sesklo and Volakas should also be of similar age (Benammi et al. [
133]; Koufos, Kostopoulos [
29]; Spassov [
11]).
Late Villafranchian. The biochronological position of Slivnitsa has been discussed in detail above. Slivnitsa should be the earliest Balkan locality, marking the beginning of the Late Villafranchian (MNQ18a), and deserves to represent a Balkan analogue of Coste San Giacomo Unit, in which unit (CSG), we have reason to place this locality. La Pietris, Romania, does not have a very clear position. The site still has
Nyctereutes megamastoides, which is absent in Olivola (but this may also be due to geographical differences). Characteristic artiodactyls are
Pliotragus ardeus (Radulescu et al. [
55]) and the cervid Rucervus (
Arvernoceros)
radulescui (Terhune et al. [
39]), which appear to be generally typical of “pre-Pachycrocuta event” faunas (Terhune et al. [
39]; Croitor et al. [
94]). The site is perhaps only slightly later than Slivnitsa and similar in age to La Puebla (Spassov [
11]), placed today (Nomade et al. [
19]) at ~2.1–2.0 Ma. Of fairly close age is probably the Romanian site Valea Graunceanului (Radulescu et al. [
55]), which has a largely similar fauna, but also
Smutsia (Pholidota) and possibly also
Pachycrocuta (?) (Terhune at al. [
39]). This site, which presents apparently wetter and forested conditions, should be later than Slivnitsa, but probably also a little later than La Pietris (Spassov [
11]), with an age perhaps corresponding to the beginning of the Olduvai subchron (?), which, however, somewhat contradicts the presence of some “pre-Pachycrocuta event” ungulates. The Geek locality of Gerakarou-1 was especially discussed in the analysis of the
Pachycrocuta event as a locality that possibly marked the first appearance of this hyena on the continent (see above). It marks the beginning of the Late Villafranchian in Greece (Koufos [
97]; Konidaris, Kostopoulos [
41,
134]) and shows many similarities with Slivnitsa. At the same time,
Pachycrocuta already occurs in Gerakarou-1, and it should be placed a little later than this Bulgarian locality (Spassov [
11]), most likely at the very beginning of the Olivola FU (MNQ18b, according to my understanding), which is why we place it here at ca. 2.1–2 Ma. Almost of the same age (~2 Ma) should be the pre-Olduvai locality of Karnezeika in Southern Greece (Sianis et al. [
95]). The fauna from the Late Villafranchian locality of Krimni-3 (Northern Greece) is placed within rather wide limits (1.8–1.5 Ma) and apparently shows the last occurrence of
Palaeotragus and
Sus strozzi s. str. in the southern Balkans, as well as the presence of the giant ostrich
Pachystruthio dmanisensis (Konidaris et al. [
135]). The presence in this fauna of
Stephanorhinus cf.
hundsheimensis contradicts this dating because this rhino is apparently known in Europe and in Anatolia from about the beginning of the Epivillafranchian (Pandolfi, Erten [
136]), and I place this locality within the indicated limits somewhat tentatively. The fauna from the Trlica cave (Montenegro), TRL11-10 level, has also been determined in similar, broad limits—1.8–1.5 Ma (Vislobokova et al. [
137]). The Greek locality of Apollonia has an important biochronological significance. It is positioned by Koufos and Kostopoulos [
29] in the Epivillafranchian. At the same time, the presence of the primitive bison
B. (
Eobison) (Sorbelli et al. [
117]) seems to confirm to one degree or another my statement (Spassov [
11]) that the site has significant similarities with Pirro (Pirro FU), which has an age of ca. 14–1.2 Ma (Napoleone et al. [
98]). However, Apollonia shows a transitional fauna to the Epivillafranchian one. In this sense, placing the locality within ~ 1.3–1.1 Ma (Konidaris, Kostopoulos [
41,
134]) or at the boundary of the Late Villafranchian s. str./Epivillafranchian, at the time of the beginning of a new strong aridification (see Spassov [
34]), i.e., at 1.2 Ma or slightly before (not later than this date) seems logical.